Carregar apresentação
A apresentação está carregando. Por favor, espere

1
21 MAI 2014 Ricardo Boavida Ferreira Sala 40 Metabolismo dos aminoácidos e proteínas. II. Anabolismo Assimilação do azoto e do enxofre (fundamentos). Biossíntese dos aminoácidos proteicos. As famílias de aminoácidos. Aminoácidos essenciais, não-essenciais e semi-essenciais. Breve referência à biossíntese dos aminoácidos raros das proteínas e dos aminoácidoa não-proteicos. Biossíntese de proteínas (fundamentos). Material de estudo: diapositivos das aulas, bibliografia recomendada e textos de apoio. LICENCIATURA EM BIOLOGIA DISCIPLINA BIOQUÍMICA Ano Lectivo de 2013/2014 Aula nº 24
. Biossíntese dos aminoácidos proteicos. As famílias de aminoácidos. Aminoácidos essenciais, não-essenciais e semi-essenciais. Breve referência à biossíntese dos aminoácidos raros das proteínas e dos aminoácidoa não-proteicos. Biossíntese de proteínas (fundamentos). Material de estudo: diapositivos das aulas, bibliografia recomendada e textos de apoio. LICENCIATURA EM BIOLOGIA DISCIPLINA BIOQUÍMICA Ano Lectivo de 2013/2014 Aula nº 24.")
2
Metabolismo dos aminoácidos e proteínas II. Anabolismo

3
ASSIMILAÇÃO DO AZOTO

4
Assimilação do carbono, do azoto e do enxofre - Definição - Quem é responsável? - Assimilação do carbono: CO 2 Hidratos de carbono - Assimilação do azoto: N 2 NH 4 + NO 3 - NO 2 - NH 4 + NH 4 + NH 4 + - Assimilação do enxofre: S 2- SO 4 2- Fotossíntese Nitrato redutase Nitrogenase Nitrito redutase Aminoácidos ESQUELETOS CARBONADOS Aminoácidos

5
reduction Nitrogen is a critical part of amino acids, nucleotides and other important biomolecules. Most prevalent form Anaerobic

6
Redução Formas de azoto inorgânico disponíveis para plantas e microrganismos autotróficos

7
Assimilação do azoto*: à semelhança da assimilação do carbono e do enxofre (e contrariamente à do fósforo), é feita por plantas e microrganismos. * Assimilação do azoto pode ser definida como o processo pelo qual o azoto passa de formas inorgânicas para combinação orgânica. Principais formas de azoto inorgânico disponíveis para plantas e microrganismos: N 2, NO 3 - e NH 4 + N 2 – Os organismos que fazem a fixação biológica do azoto contêm um compexo multienzimático – a nitrogenase – capaz de quebrar a ligação covalente tripla que une os dois átomos de azoto do N 2. Nas plantas da família das Leguminosas, estabelece-se uma relação simbionte entre bactéria fixadora do azoto (do género Rhizobium), que fornece NH 4 + à planta, e a planta, que abastece a bactéria de fotoassimilados (i.e., hidratos de carbono). A nitrogenase é inibida pelo O 2, o que justifica a presença da legoglobina nos nódulos, que ficam, assim, avermelhados. A parte proteica da legoglobina é fornecida pela planta e o respectivo grupo heme pela bactéria simbionte. NO 3 - - Trata-se de uma forma de azoto que pode ser absorvida e armazenada nos vacúolos das folhas das plantas. A sua redução ao nível de amoníaco dá-se pela acção sequencial de duas importantes enzimas, que consomem potencial redutor produzido pelas reacções fotoquímicas da fotossíntese: a nitrato redutase (NR), que reduz o nitrato a nitrito, e a nitrito redutase (NiR), que reduz o nitrito a amoníaco. NH 4 + - Trata-se de uma forma tóxica de azoto, não podendo, por isso, acumular-se nas células (desacopla as cadeias de transporte de electrões). O amónio é assimilado, isto é, incorporado em aminoácidos pelo funcionamento do ciclo da glutamato sintase.
, que fornece NH 4 + à planta, e a planta, que abastece a bactéria de fotoassimilados (i.e., hidratos de carbono). A nitrogenase é inibida pelo O 2, o que justifica a presença da legoglobina nos nódulos, que ficam, assim, avermelhados. A parte proteica da legoglobina é fornecida pela planta e o respectivo grupo heme pela bactéria simbionte. NO Trata-se de uma forma de azoto que pode ser absorvida e armazenada nos vacúolos das folhas das plantas. A sua redução ao nível de amoníaco dá-se pela acção sequencial de duas importantes enzimas, que consomem potencial redutor produzido pelas reacções fotoquímicas da fotossíntese: a nitrato redutase (NR), que reduz o nitrato a nitrito, e a nitrito redutase (NiR), que reduz o nitrito a amoníaco. NH Trata-se de uma forma tóxica de azoto, não podendo, por isso, acumular-se nas células (desacopla as cadeias de transporte de electrões). O amónio é assimilado, isto é, incorporado em aminoácidos pelo funcionamento do ciclo da glutamato sintase..")
8
Nitrogen fixation

9
Fixation of atmospheric N 2 Certain free living bacteria Symbiotic bacteria Non-enzymatic (Haber method, 500 o C, hundreds of Atms) N 2 + 3H 2 2NH 3 Enzymatic (nitrogenase complex, highly conserved) N 2 + 10H + + 8e - + 16ATP 2NH 4 + + 16ADP + 16P i + H 2 1 e - /2ATP per cycle 2ATP binding shifts reduction potential Fe-S clusters Iron Molybdenum 8 electrons (6 for N 2, 2 for H 2 )
 N 2 + 3H 2 2NH 3 Enzymatic (nitrogenase complex, highly conserved) N H + + 8e ATP 2NH ADP + 16P i + H 2 1 e - /2ATP per cycle 2ATP binding shifts reduction potential Fe-S clusters Iron Molybdenum 8 electrons (6 for N 2, 2 for H 2 )")
10
ADP, Fe, S, MoFe Iron-molybdenum cofactor Nitrogenase Oxygen toxicity to bacterial nitrogenase: plant derived leghemoglobin Mo, Fe S homocitrate

11
Reduced nitrogen (NH 4 + ) via Glu and Gln Gln synthetase (two step; key regulatory) γ-Glu phosphate (mixed anhydride) γ-Glu phosphate + NH 4 + = Gln Ala, Gly and others are partial/allosteric I Primary regulatory point Adenylation, covalent, inhibitory Adenylyltransferase, AT Gln synthetase
 via Glu and Gln Gln synthetase (two step; key regulatory) γ-Glu phosphate (mixed anhydride) γ-Glu phosphate + NH 4 + = Gln Ala, Gly and others are partial/allosteric I Primary regulatory point Adenylation, covalent, inhibitory Adenylyltransferase, AT Gln synthetase")
12
Subunit structure of Gln synthetase (12 identical subunits, active sites) Side view
 Side view")
13
Top view

14
Uridylylation of Tyr (P II -UMP stimulates deadenylation) AT = adenyltransferase
 AT = adenyltransferase")
15
Biossíntese dos aminoácidos Vias de assimilação do amónio Aminoácido Esqueleto carbonado Amónio +

16
H 2 N – C COOH | | R H Biossíntese dos aminoácidos Esqueleto carbonado: Fornecido por metabolitos das seguintes vias metabólicas: - Glicólise; - Via dos fosfatos de pentose; - Ciclo do ácido cítrico. Vias de assimilação do amónio Nitrogenase N 2 (atmosfera) NH 4 + NH 4 + (solo) NO 2 - NO 3 - (solo) Nitrito redutase Nitrato redutase
 NH 4 + NH 4 + (solo) NO 2 - NO 3 - (solo) Nitrito redutase Nitrato redutase.")
17
A via GS/GOGAT consome ATP mas consegue assimilar o NH 3 eficientemente quando a sua concentração é baixa – isto porque o valor do K m da enzima GS para o NH 3 é muito mais baixo que o da enzima GDH. Foi, por isso, sugerido que o ATP gasto pela via GS/GOGAT é o preço que as células que possuem esta via têm de pagar para conseguirem assimilar o NH 3 presente em baixas concentrações.

18
Ammonia Assimilation and Recycling The glutamate synthase or GS-GOGAT cycle For many years it was thought that bacteria and higher plants assimilate ammonia into glutamate via the GDH pathway, as in certain fungi and yeasts. However, in bacteria it became clear in 1970 that an alternative pathway of ammonia assimilation [involving glutamine synthetase (GS) [EC 6.3.1.2] and an NADPH-dependent glutamine:2-oxoglutarate amidotransferase (GOGAT) [EC 1.4.1.13] or glutamate synthase, must be operating when ammonia is present in the growth medium at low levels. Thus, N-starvation leads to derepression and activation of GS (with a high affinity for NH 3 ) and derepression of GOGAT, and repression of GDH (with a relatively low affinity for NH 3 ). High ammonia availability leads to repression and deactivation of GS and induction of GDH. GDH NH 3 + 2-oxoglutarate + NADPH + H + glutamate + NADP + GS-GOGAT NH 3 + glutamate + ATP ---> glutamine + ADP + P i glutamine + 2-oxoglutarate + NADPH + H + ---> 2 glutamate + NADP + Both the GDH and GS-GOGAT pathways produce 1 mole of glutamate from 1 mole each of NH 3, 2-oxoglutarate and NADPH. But note that the GS-GOGAT pathway is energetically more costly than the GDH pathway, consuming 1 ATP. Escherichia coli is now known to have two primary pathways for glutamate synthesis. The GS-GOGAT pathway is essential for glutamate synthesis at low ammonium concentrations and for regulation of the glutamine pool, and is used when the cell is not under energy limitation. The GDH pathway is used in glutamate synthesis when the cell is limited for energy (and carbon; i.e. glucose-limited growth) but ammonium and phosphate are present in excess. Synechocystis sp. strain PCC 6803 utilizes the GS-GOGAT pathway as the primary pathway of ammonia assimilation, but the presence of GDH appears to offer a selective advantage for the cyanobacterium under nonexponential growth conditions. These dual pathways may be common to bacteria, cyanobacteria, algae, yeasts and fungi. Re-examination of ammonia assimilation in yeasts and fungi now reveals the operation of alternative pathways of glutamate synthesis, independent of NADPH-GDH:
 [EC ] and an NADPH-dependent glutamine:2-oxoglutarate amidotransferase (GOGAT) [EC ] or glutamate synthase, must be operating when ammonia is present in the growth medium at low levels. Thus, N-starvation leads to derepression and activation of GS (with a high affinity for NH 3 ) and derepression of GOGAT, and repression of GDH (with a relatively low affinity for NH 3 ). High ammonia availability leads to repression and deactivation of GS and induction of GDH. GDH NH oxoglutarate + NADPH + H + glutamate + NADP + GS-GOGAT NH 3 + glutamate + ATP ---> glutamine + ADP + P i glutamine + 2-oxoglutarate + NADPH + H + ---> 2 glutamate + NADP + Both the GDH and GS-GOGAT pathways produce 1 mole of glutamate from 1 mole each of NH 3, 2-oxoglutarate and NADPH. But note that the GS-GOGAT pathway is energetically more costly than the GDH pathway, consuming 1 ATP. Escherichia coli is now known to have two primary pathways for glutamate synthesis. The GS-GOGAT pathway is essential for glutamate synthesis at low ammonium concentrations and for regulation of the glutamine pool, and is used when the cell is not under energy limitation. The GDH pathway is used in glutamate synthesis when the cell is limited for energy (and carbon; i.e. glucose-limited growth) but ammonium and phosphate are present in excess. Synechocystis sp. strain PCC 6803 utilizes the GS-GOGAT pathway as the primary pathway of ammonia assimilation, but the presence of GDH appears to offer a selective advantage for the cyanobacterium under nonexponential growth conditions. These dual pathways may be common to bacteria, cyanobacteria, algae, yeasts and fungi. Re-examination of ammonia assimilation in yeasts and fungi now reveals the operation of alternative pathways of glutamate synthesis, independent of NADPH-GDH:.")
19
Mutants of Aspergillus nidulans lacking NADP-GDH activity grow more poorly than wild-type strains on ammonium as a sole nitrogen source. The leaky growth of these mutants is indicative of an alternative pathway of ammonium assimilation and glutamate biosynthesis. A. nidulans mutants disrupted in the gltA encoding GOGAT, were found to be dispensable for growth on ammonium in the presence of NADP-GDH. However, a strain carrying the gltA inactivation together with an NADP-GDH structural gene mutation (gdhA) was unable to grow on ammonium or on nitrogen sources metabolized via ammonium. Schizosaccharomyces pombe mutants lacking either NADPH-GDH or GOGAT are still able to grow on ammonium as sole nitrogen source. Complete lack of growth on ammonium as sole N source is seen only in double mutants lacking both NADPH-GDH and GOGAT. In contrast to Candida utilis, analysis of 15 N-ammonium assimilation in actively growing mycelium of Agaricus bisporus indicates participation of the GS-GOGAT pathway, and no participation of NADP-GDH. 13 NH 3 tracer studies indicate that the GS-GOGAT pathway is the major route of ammonium assimilation in Candida albicans and also in nitrogen-starved cultures of Saccharomyces cerevisiae and Candida tropicalis. The yeast Saccharomyces cerevisiae synthesizes glutamate through the action of either NADP-glutamate dehydrogenase (NADP-GDH), encoded by GDH1 (under conditions of ammonia excess), or through the combined action of glutamine synthetase (GS) and glutamate synthase (GOGAT), encoded by GLN1 and GLT1 (under conditions of ammonia limitation). Dynamic modeling indicates that the GS-GOGAT pathway plays a more important physiological role in yeast than is generally assumed. However, a double mutant of S. cerevisiae lacking NADP-GDH and GOGAT activities was able to grow on ammonium as the sole nitrogen source and thus to synthesize glutamate through a third pathway. A computer search for similarities between the GDH1 nucleotide sequence and the complete yeast genome led to the discovery that GDH1 showed high identity to an open reading frame (GDH3) on chromosome I. Triple mutants impaired in GDH1, GLT1, and GDH3 are strict glutamate auxotrophs, indicating that GDH3 plays a significant physiological role, providing glutamate when GDH1 and GLT1 are impaired. This appears to be the first example of a microorganism possessing three pathways for glutamate biosynthesis. Following the discovery of glutamate synthase (GOGAT) in bacteria, a similar activity was sought in plants. A ferredoxin-dependent glutamate synthase [EC 1.4.7.1] was discovered in photosynthetic tissues of higher plants in 1974, and an NADH-dependent "glutamate synthetase" in non-photosynthetic plant tissues in the same year.
 was unable to grow on ammonium or on nitrogen sources metabolized via ammonium. Schizosaccharomyces pombe mutants lacking either NADPH-GDH or GOGAT are still able to grow on ammonium as sole nitrogen source. Complete lack of growth on ammonium as sole N source is seen only in double mutants lacking both NADPH-GDH and GOGAT. In contrast to Candida utilis, analysis of 15 N-ammonium assimilation in actively growing mycelium of Agaricus bisporus indicates participation of the GS-GOGAT pathway, and no participation of NADP-GDH. 13 NH 3 tracer studies indicate that the GS-GOGAT pathway is the major route of ammonium assimilation in Candida albicans and also in nitrogen-starved cultures of Saccharomyces cerevisiae and Candida tropicalis. The yeast Saccharomyces cerevisiae synthesizes glutamate through the action of either NADP-glutamate dehydrogenase (NADP-GDH), encoded by GDH1 (under conditions of ammonia excess), or through the combined action of glutamine synthetase (GS) and glutamate synthase (GOGAT), encoded by GLN1 and GLT1 (under conditions of ammonia limitation). Dynamic modeling indicates that the GS-GOGAT pathway plays a more important physiological role in yeast than is generally assumed. However, a double mutant of S. cerevisiae lacking NADP-GDH and GOGAT activities was able to grow on ammonium as the sole nitrogen source and thus to synthesize glutamate through a third pathway. A computer search for similarities between the GDH1 nucleotide sequence and the complete yeast genome led to the discovery that GDH1 showed high identity to an open reading frame (GDH3) on chromosome I. Triple mutants impaired in GDH1, GLT1, and GDH3 are strict glutamate auxotrophs, indicating that GDH3 plays a significant physiological role, providing glutamate when GDH1 and GLT1 are impaired. This appears to be the first example of a microorganism possessing three pathways for glutamate biosynthesis. Following the discovery of glutamate synthase (GOGAT) in bacteria, a similar activity was sought in plants. A ferredoxin-dependent glutamate synthase [EC ] was discovered in photosynthetic tissues of higher plants in 1974, and an NADH-dependent glutamate synthetase in non-photosynthetic plant tissues in the same year..")
20
O ciclo da glutamato sintase

21
Evidence in favor of the operation of the GS-GOGAT cycle as the primary pathway of ammonia assimilation in higher plants includes: - Almost complete inhibition of 15 NH 4 + assimilation by the glutamine synthetase (GS) inhibitor, methionine sulfoximine (MSX). - Quantitative analysis of 15 NH 4 + in Lemna minor is consistent with incorporation of 15 N primarily into glutamine- amide, followed by transfer to glutamate and the amino-group of glutamine via the action of GOGAT and GS, respectively, provided that it is assumed that the GS-GOGAT cycle is compartmentilized in the chloroplast, and that a second site of glutamine synthesis occurs in the cytoplasm. - The maize gdh1-null mutant exhibits about 5% of the total GDH enzyme activity of wildtype plants. Although this mutant exhibits a slightly reduced total rate of 15 NH 4 + assimilation, when methionine sulfoximine (MSX), a potent inhibitor of GS is supplied, this completely blocks 15 NH 4 + assimilation in both the mutant and wildtype roots and shoots. The contribution of GDH to net ammonia assimilation is small in comparison to that catalyzed by the GS-GOGAT cycle. - Mutants of Arabidopsis and barley defective in GS or GOGAT exhibit markedly impaired ammonia assimilation, especially under photorespiratory conditions. Enzyme kinetic considerations also suggest a role for the GS-GOGAT pathway in ammonia assimilation at low tissue/cell ammonia concentrations. GS has a much higher affinity for ammonia than GDH and is viewed as a scavenger of ammonia in bacteria and in plants. The major role of GDH in tissue cultured cells is the oxidation of glutamate to provide sufficient carbon skeletons for effective functioning of the TCA cycle. In wildtype Arapidopsis, GDH1 mRNA accumulates to high levels in dark-adapted or sucrose-starved plants; light or sucrose treatment each repress GDH1 mRNA accumulation. These results suggest that the GDH1 gene product functions in the direction of glutamate catabolism under carbon-limiting conditions. Low levels of GDH1 mRNA present in leaves of light-grown plants can be induced by exogenously supplied ammonia. Under such conditions of carbon and ammonia excess, GDH1 may function in the direction of glutamate biosynthesis. The recessive Arabidopsis glutamate dehydrogenase-deficient mutant allele gdh1-1 cosegregates with the GDH1. The gdh1-1 mutant displays a conditional phenotype; seedling growth is specifically retarded on media containing exogenously supplied inorganic nitrogen, suggesting that GDH1 plays a nonredundant role in ammonia assimilation under conditions of inorganic nitrogen excess. This is consistent with the fact that the levels of mRNA for GDH1 and chloroplastic glutamine synthetase (GS2) are reciprocally regulated by light.
, a potent inhibitor of GS is supplied, this completely blocks 15 NH 4 + assimilation in both the mutant and wildtype roots and shoots. The contribution of GDH to net ammonia assimilation is small in comparison to that catalyzed by the GS-GOGAT cycle. - Mutants of Arabidopsis and barley defective in GS or GOGAT exhibit markedly impaired ammonia assimilation, especially under photorespiratory conditions. Enzyme kinetic considerations also suggest a role for the GS-GOGAT pathway in ammonia assimilation at low tissue/cell ammonia concentrations. GS has a much higher affinity for ammonia than GDH and is viewed as a scavenger of ammonia in bacteria and in plants. The major role of GDH in tissue cultured cells is the oxidation of glutamate to provide sufficient carbon skeletons for effective functioning of the TCA cycle. In wildtype Arapidopsis, GDH1 mRNA accumulates to high levels in dark-adapted or sucrose-starved plants; light or sucrose treatment each repress GDH1 mRNA accumulation. These results suggest that the GDH1 gene product functions in the direction of glutamate catabolism under carbon-limiting conditions. Low levels of GDH1 mRNA present in leaves of light-grown plants can be induced by exogenously supplied ammonia. Under such conditions of carbon and ammonia excess, GDH1 may function in the direction of glutamate biosynthesis. The recessive Arabidopsis glutamate dehydrogenase-deficient mutant allele gdh1-1 cosegregates with the GDH1. The gdh1-1 mutant displays a conditional phenotype; seedling growth is specifically retarded on media containing exogenously supplied inorganic nitrogen, suggesting that GDH1 plays a nonredundant role in ammonia assimilation under conditions of inorganic nitrogen excess. This is consistent with the fact that the levels of mRNA for GDH1 and chloroplastic glutamine synthetase (GS2) are reciprocally regulated by light..")
22
Aminoácidos essenciais Os organismos diferem muito na sua capacidade de sintetizarem os aminoácidos de que necessitam para a síntese proteica. As plantas autotróficas e muitos microrganismos são auto-suficientes, no sentido em que conseguem sintetizar os 20 aminoácidos proteicos. A bactéria Leuconostoc mesenteroides consegue apenas sintetizar quatro aminoácidos proteicos, enquanto que a bactéria Lactobacillus, que cresce no leite, não consegue sintetizar nenhum. Os mamíferos estão numa posição intermédia, sendo capazes de sintetizar metade dos aminoácidos proteicos – o que significa que têm de obter os outros na dieta alimentar. Os aminoácidos que não podem ser sintetizados por um organismo em quantidade suficiente são denominados aminoácidos essenciais ou indispensáveis e têm que ser fornecidos pela dieta alimentar. Aqueles que podem ser sintetizados pelo organismo, a partir de precursores disponíveis, em quantidade suficiente para satisfazer as suas necessidades são denominados não-essenciais ou dispensáveis. São considerados aminoácidos essenciais para o homem a valina (Val), a leucina, (Leu), a isoloeucina (Ile), o triptofano (Trp), a fenilalanina (Phe), a lisina (Lys), a metionina (Met) e a treonina (Thr). A arginina (Arg) e a histidina (His) são sintetizados em quantidade suficiente para satisfazer as necessidades do homem adulto, mas não as de uma criança em crescimento. Estes aminoácidos são, por isso, designados por semi- ou meio-essenciais. A tirosina (Tyr) e a cisteína (Cys) são considerados não-essenciais se a dieta alimentar tiver quantidades suficientes de fenilalanina e de metionina, respectivamente. Isto porque os mamíferos formam tirosina directamente a partir da fenilalanina e porque a cisteína deriva o seu enxofre da metionina.
, a leucina, (Leu), a isoloeucina (Ile), o triptofano (Trp), a fenilalanina (Phe), a lisina (Lys), a metionina (Met) e a treonina (Thr). A arginina (Arg) e a histidina (His) são sintetizados em quantidade suficiente para satisfazer as necessidades do homem adulto, mas não as de uma criança em crescimento. Estes aminoácidos são, por isso, designados por semi- ou meio-essenciais. A tirosina (Tyr) e a cisteína (Cys) são considerados não-essenciais se a dieta alimentar tiver quantidades suficientes de fenilalanina e de metionina, respectivamente. Isto porque os mamíferos formam tirosina directamente a partir da fenilalanina e porque a cisteína deriva o seu enxofre da metionina..")
23
Em geral, os aminoácidos essenciais são aqueles com estruturas mais complicadas e formados por vias metabólicas mais complexas, enquanto que os aminoácidos não-essenciais têm biossínteses mais simples, a partir de precursores que estão normalmente presentes em todas as células. A deficiência em um ou mais aminoácidos essenciais na dieta de um organismo origina, tipicamente, um balanço de azoto negativo, isto é, o azoto total excretado pelo organismo excede o que é absorvido, indicando degradação das proteínas dos tecidos para fornecer o (ou os) aminoácido que falta para a síntese de novas proteínas prioritárias ou essenciais à sobrevivência do organismo. Os restantes aminoácidos que compôem essas proteínas acumulam-se e sofrem catabolismo – daí a excreção do azoto e o balanço de azoto negativo.
 aminoácido que falta para a síntese de novas proteínas prioritárias ou essenciais à sobrevivência do organismo. Os restantes aminoácidos que compôem essas proteínas acumulam-se e sofrem catabolismo – daí a excreção do azoto e o balanço de azoto negativo..")
24
BIOSSÍNTESE DOS AMINOÁCIDOS

25
5-phosphoribosyl-1-pyrophosphate (PRPP) from ribose phosphate pyrophokinase: new intermediate in amino acid and nucleotide synthesis
 from ribose phosphate pyrophokinase: new intermediate in amino acid and nucleotide synthesis")
26
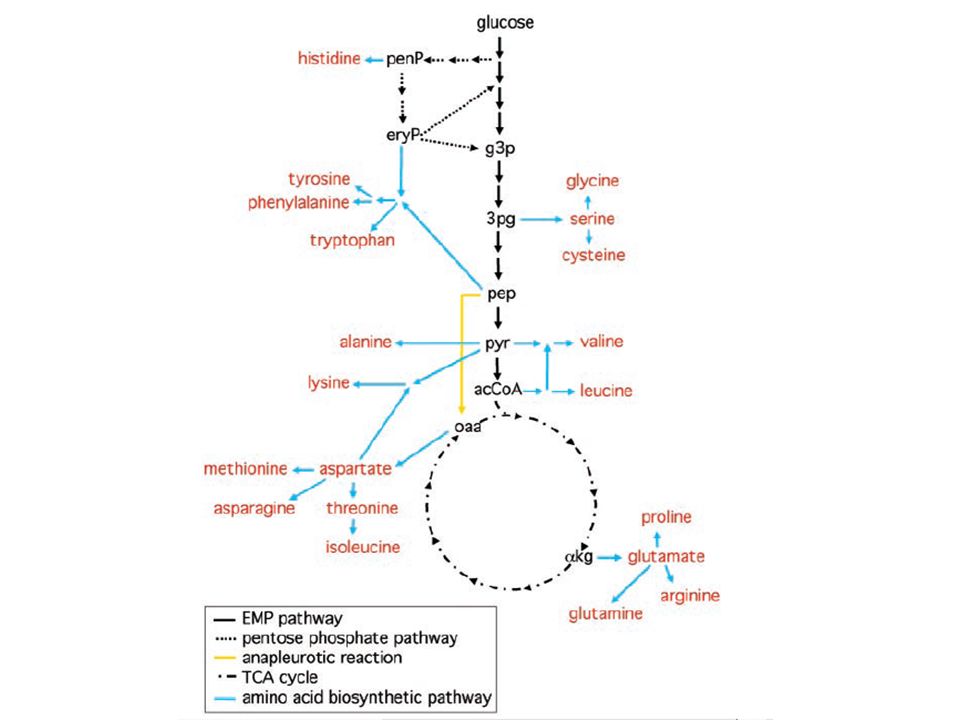
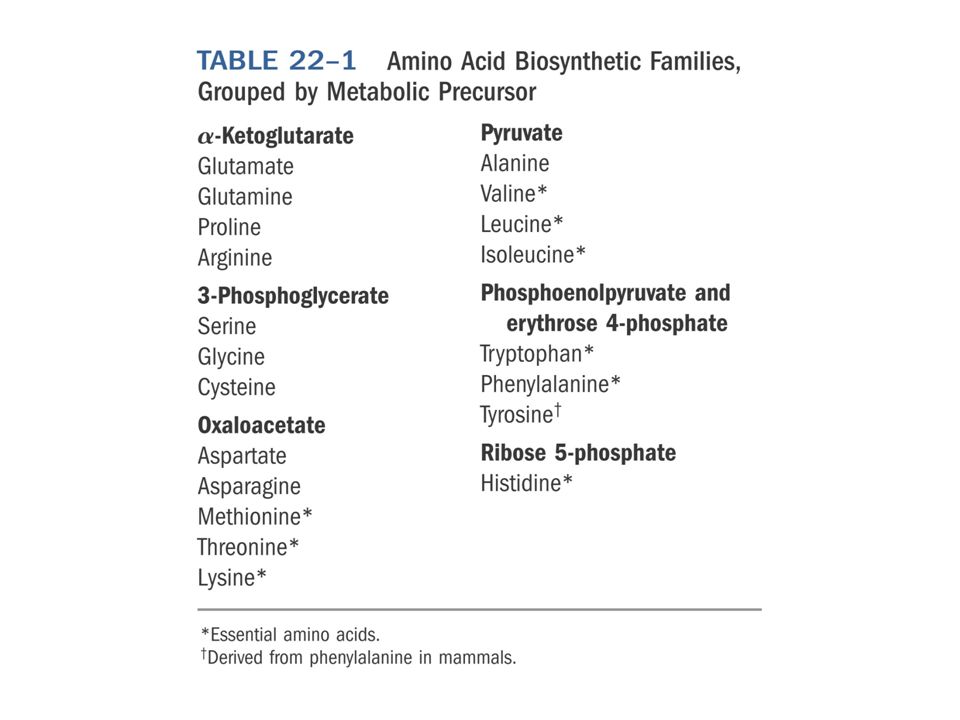
Os 20 aminoácidos proteicos são agrupados em 6 famílias de acordo com os metabolitos que lhes fornecem o esqueleto carbonado:

27
Metabolic precursor organization Glycolytic, citric acid and pentose phosphate intermediates are the carbon skeleton sources Glutamine and Glutamate are the N sources 20 common amino acid pathways (bacterial)
")
30
Biosynthetic Families Metabolic PrecursorsAmino Acids α-Ketoglutarate Glutamate, Glutamine, Proline, Arginine 3-Phosphoglycerate Serine, Glycine, Cysteine Oxaloacetate Aspartate, Asparagine, Methionine, Threonine, Lysine Pyruvate Alanine, Valine, Leucine, Isoleucine Phosphoenolpyruvate Tryptophan, Phenlyalanine, Tyrosine and erythrose 4-phosphate Ribose 5-phosphate Histidine

31
Ketoglutarate Glutamate GlutamineProlineArginine The α-Ketoglutarate or Glutamate Family O 2-oxoglutarato, um intermediário do ciclo do ácido cítrico, é o precursor da síntese do glutamato e dos outros membros desta família, a glutamina, a prolina, a arginina e, nos fungos e em Euglena, a lisina. Proline is cyclized Glutamate

32
Arg pathway missing in mammals

33
The 3-Phosphoglycerate or Serine Family 3- Phosphoglycerate Serine GlycineCysteine O 3-fosfoglicerato, um intermediário da glicólise, é o precursor dos aminoácidos da família da serina, que incluem, além deste aminoácido, a glicina e a cisteína.

34
Biosynthesis of serine/glycine

35
Biosynthesis of cysteine from serine in bacteria and plants: A via da sulfidrilação directa

36
Biosynthesis of cysteine from homocysteine and serine in mammals: A via da transulfuração Mammals: methionine and serine

37
37 Assimilação do enxofre na natureza: é feita por plantas e microrganismos A assimilação do enxofre na natureza, isto é, a incorporação do enxofre inorgânico no aminoácido cisteína, é feita por plantas e microrganismos. Há duas vias para a síntese de cisteína nos seres vivos: - A via da sulfidrilação directa, em que plantas e microrganismos usam H 2 S para sintetizar cisteína; - A via da transulfuração, na qual os mamíferos utilizam o esqueleto carbonado da serina e o enxofre da metionina para formar cisteína. Por este motivo, a metionina, mas não a cisteína, é um aminoácido essencial para os mamíferos, homem incluído.

38
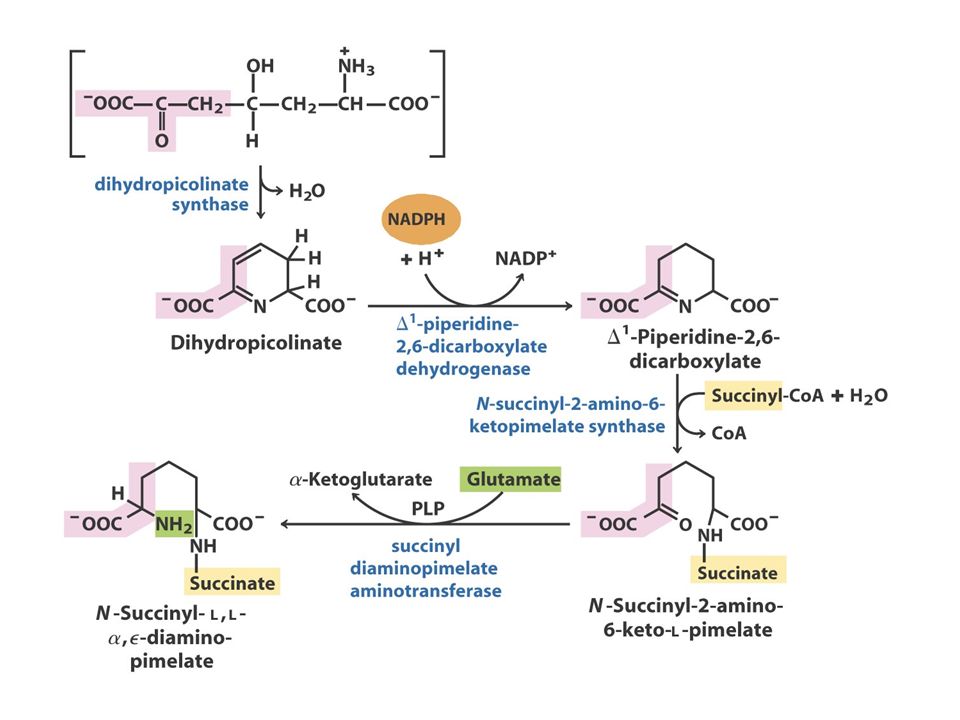
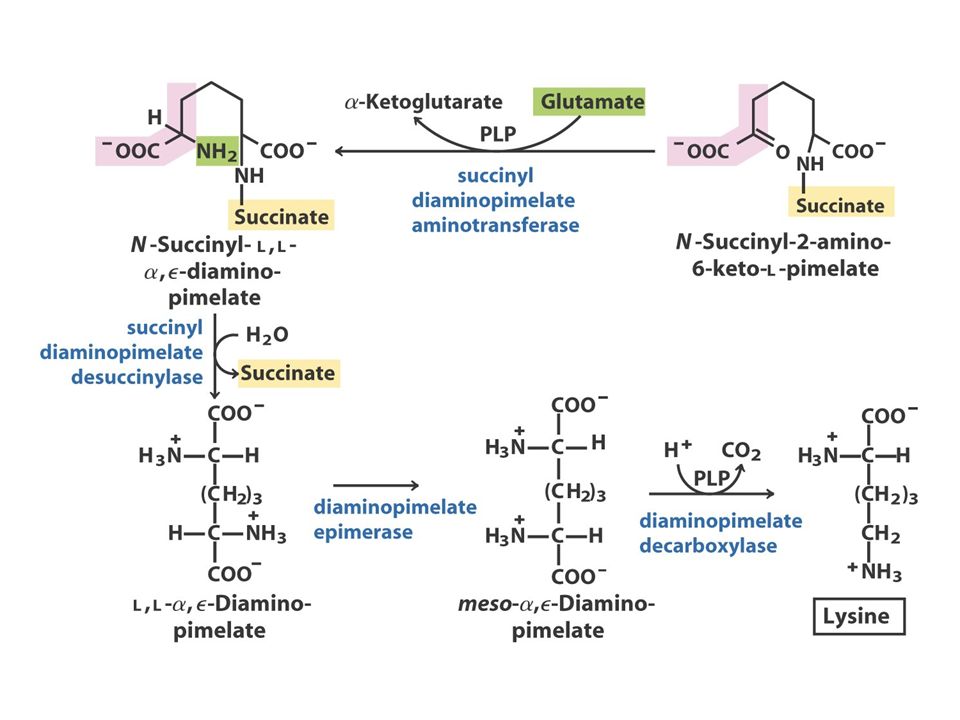
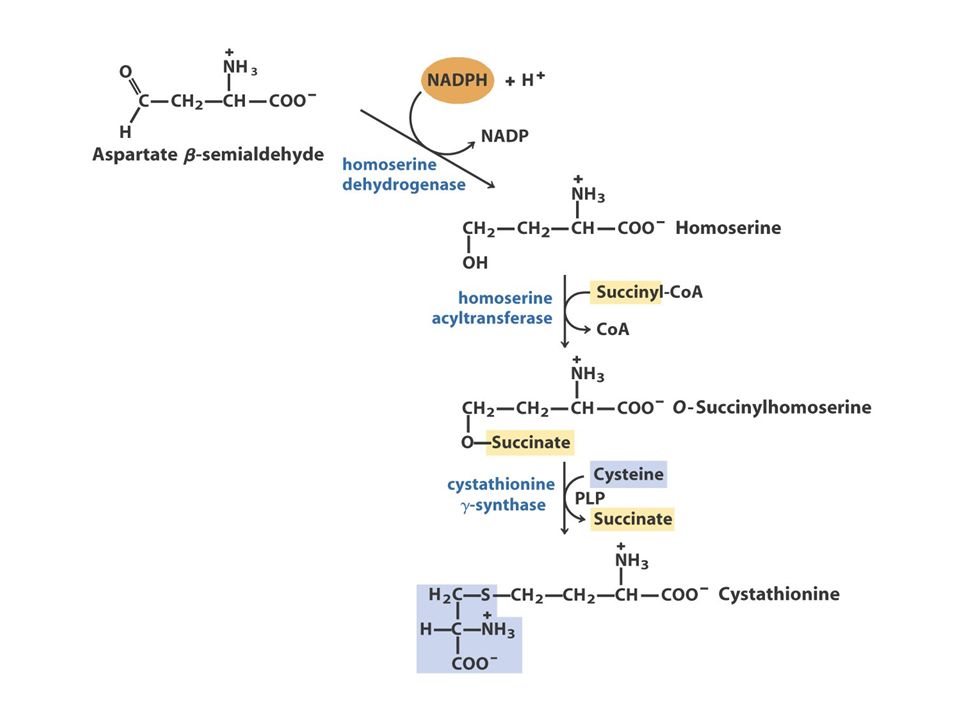
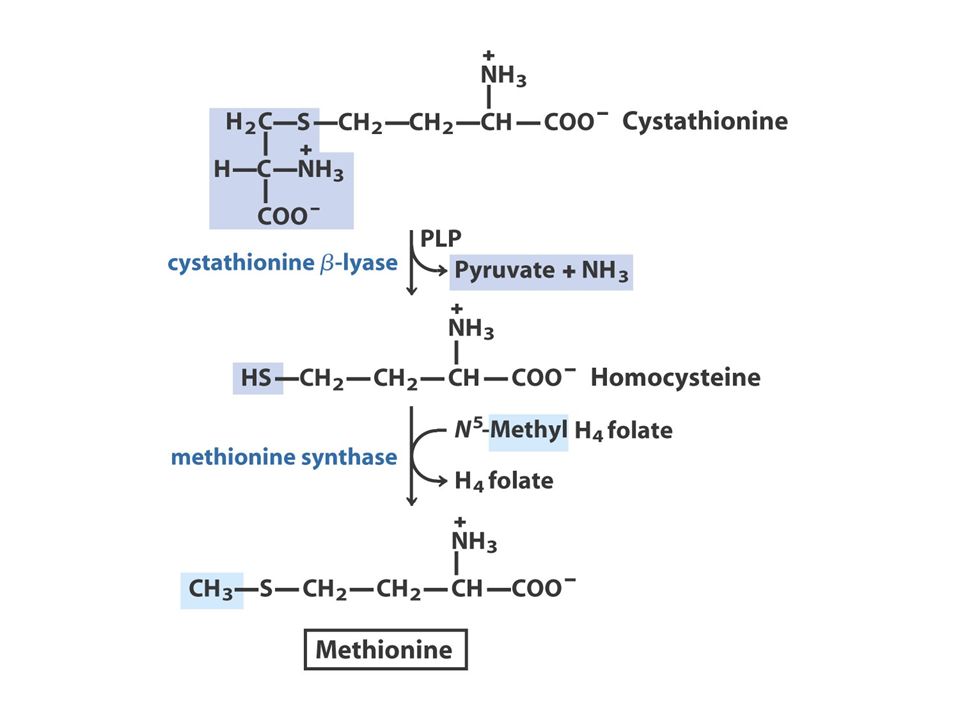
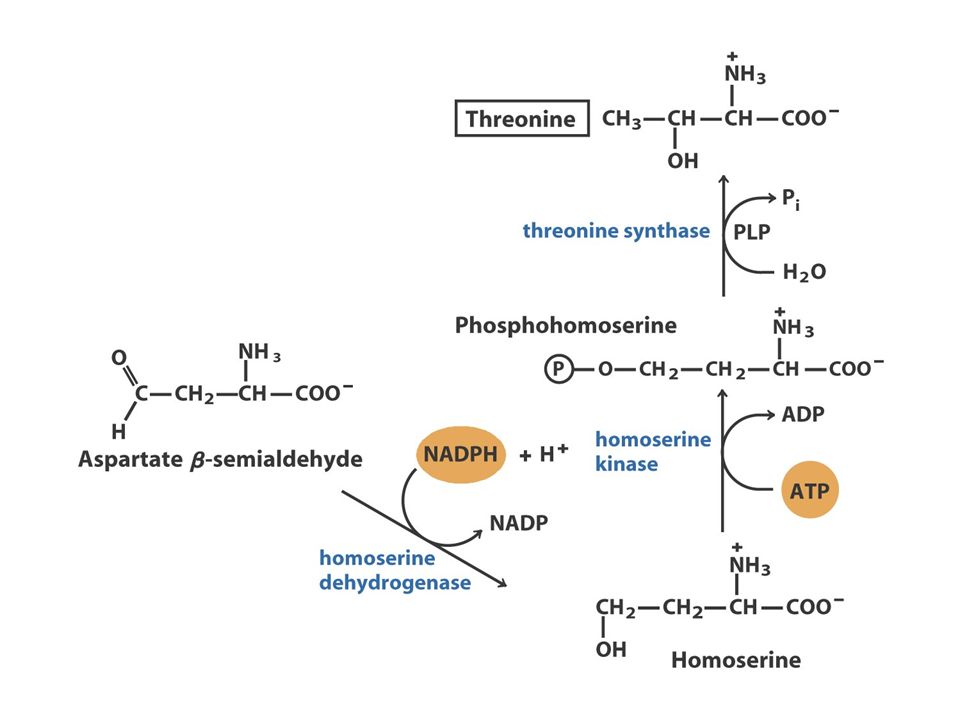
The Oxaloacetate or Aspartate Family Oxaloacetate Aspartate AsparagineMethionineLysineThreonine O oxaloacetato, um intermediário do ciclo do ácido cítrico, fornece o esqueleto carbonado para a síntese dos seis aminoácidos da família do aspartato: aspartato, asparagina, lisina (em bactérias e plantas, mas não em fungos), metionina, treonina e isoleucina. A isoleucina é, muitas vezes, incluída na família do piruvato, porque quatro das cinco enzimas que participam na sua biossíntese são comuns às enzimas da via de síntese da valina. A metionina obtém o seu enxofre a partir da cisteína.

39
Aspartate to Lysine: 10 step synthesis

45
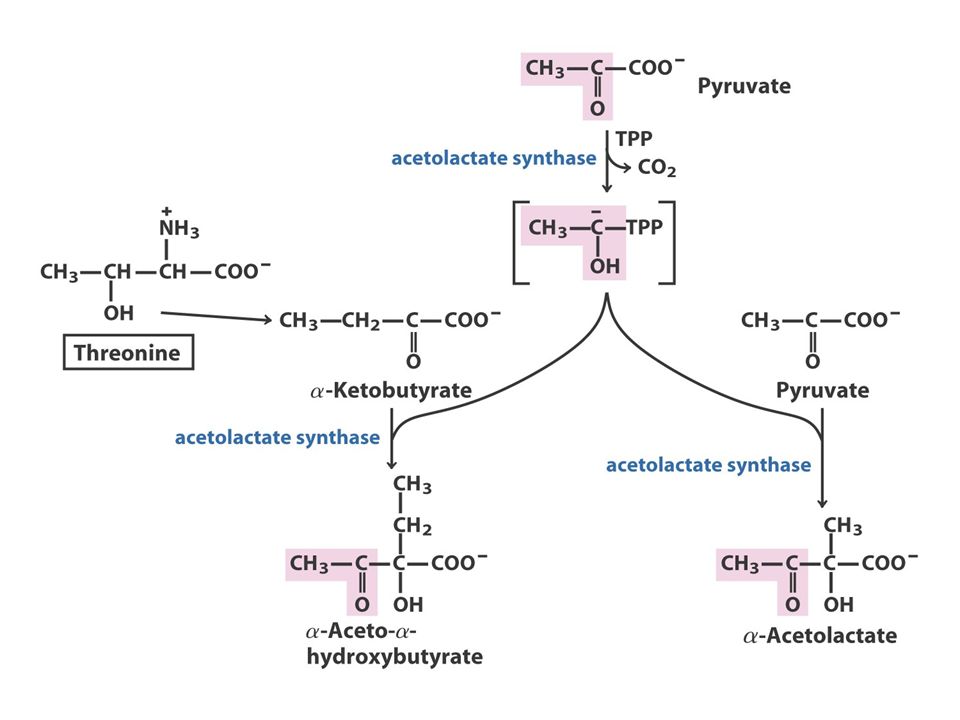
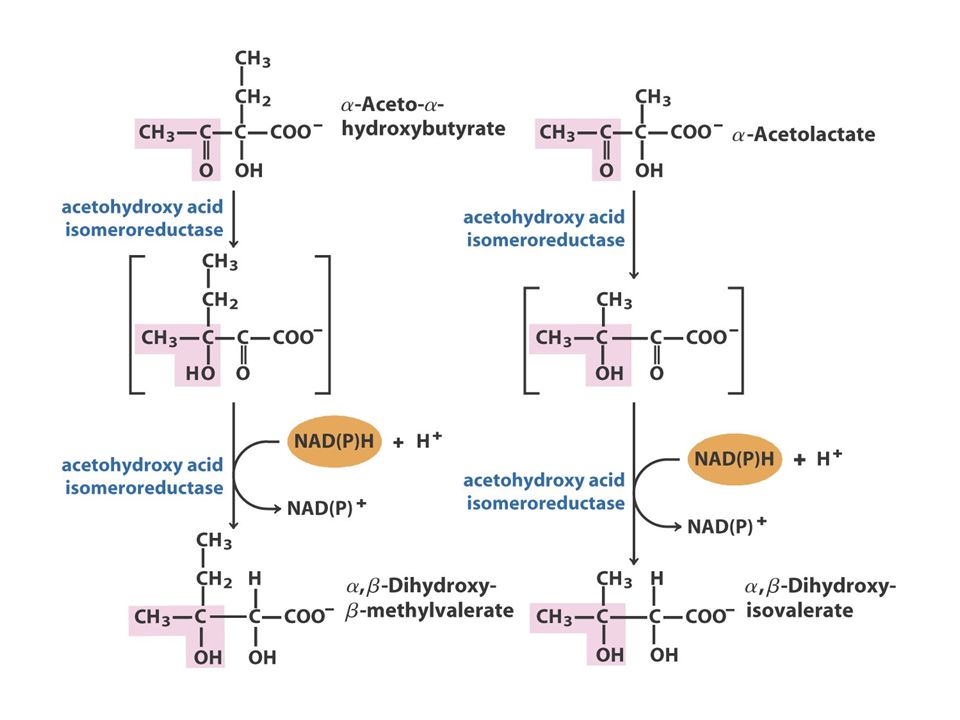
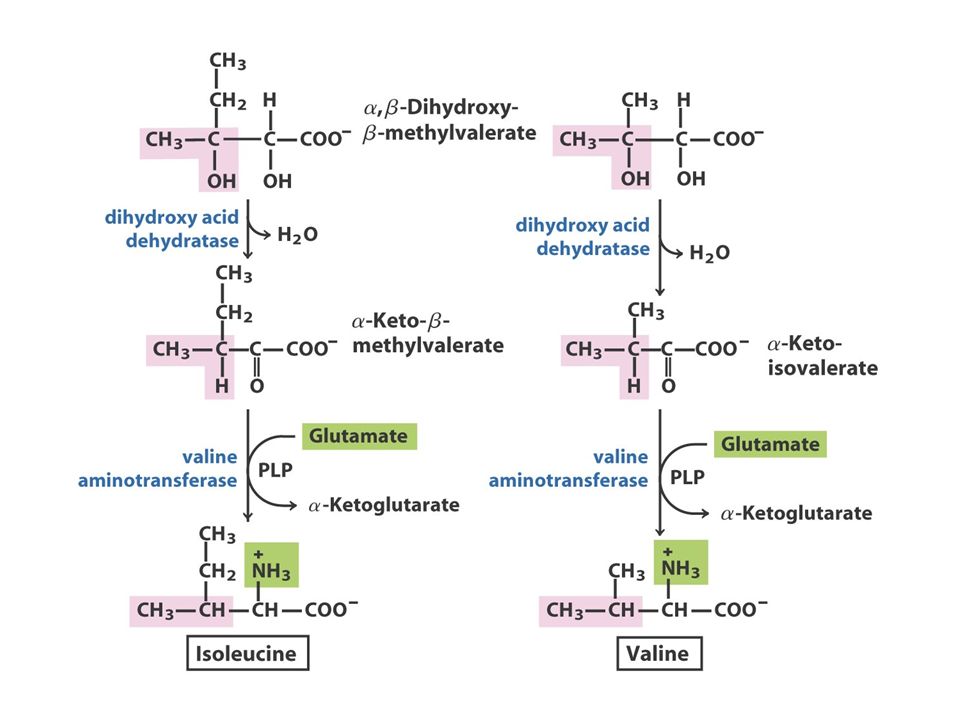
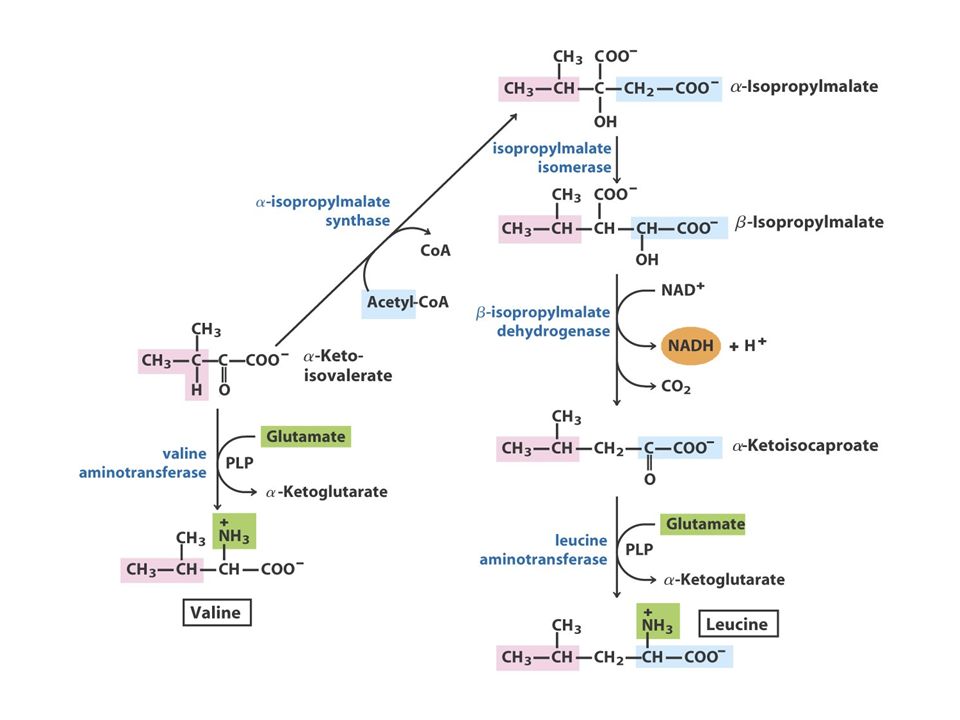
The Pyruvate Family Pyruvate AlanineValineLeucineIsoleucine O piruvato, o produto final da glicólise, fornece os esqueletos carbonados para a síntese da alanina e valina e quatro dos seis átomos de carbono da leucina. Além disso, o piruvato contribui com dois carbonos para a síntese da isoleucina e, em média, com 2,5 átomos para a síntese da lisina nas plantas e bactérias. Como referido anteriormente, a isoleucina, um membro da família do aspartato, é frequentemente incluída na família do piruvato.

50
The Phosphoenolpyruvate and Erythrose 4- phosphate or Aromatic Family Phosphoenolpyruvate + erythrose 4-phosphate Chorismate TryptophanTyrosinePhenylalanine Fenilalanina, tirosina e triptofano são sintetizados a partir do fosfoenolpiruvato, um intermediário da glicólise, e da eritrose-4-fosfato, um intermediário da via dos fosfatos de pentose. Estes aminoácidos são sintetizados por uma via metabólica ramificada divergente, em que o corismato é o principal ponto de ramificação.

51
Chorismate: intermediate in aromatic amino acid biosynthesis in bacteria and plants Carbon from erythrose 4 phosphate Carbon from phosphoenolpyruvate Via do chiquimato: Permite a síntese de um anel benzénico (aromático) O corismato é sintetizado por uma via composta por sete reacções, conhecida por via do chiquimato, onde é construído um anel de benzeno. EPSPS

52
Biosynthesis of Trp from chorismate in bacteria and plants

53
Biosynthesis of Phe and Tyr from chorismate in bacteria and plants involves prephenate Nalguns organismos, incluindo o homem, a tirosina pode ser sintetizada por hidroxilação da fenilalanina, numa reacção irreversível catalisada pela fenilalanina hidroxilase. Isto explica que a tirosina não seja um aminoácido essencial para o homem, uma vez que pode ser formada a partir da fenilalanina que ingerimos na dieta alimentar. Mas a fenilalanina é um aminoácido essencial porque a reacção catalisada pela fenilalanina hidroxilase é irreversível, isto é, não podemos formar fenilalanina a partir da tirosina. Animals (Phe hydroxylation)
.")
54
54 O glifosato, um herbicida de grande sucesso Glyphosate (N-(phosphonomethyl)glycine) is a non-selective systemic herbicide, absorbed through the leaves, used to kill weeds, especially perennials. Some crops have been genetically engineered to be resistant to it. Glyphosate was first sold by Monsanto under the tradename Roundup, but is no longer under patent. Glyphosate is an aminophosphonic analogue of the natural amino acid glycine and the name is a contraction of glycine, phospho-, and -ate. Glyphosate was first discovered to have herbicidal activity in 1970 by John Franz, while working for Monsanto. Some microorganisms have a version of 5-enolpyruvoyl-shikimate-3-phosphate synthetase (EPSPS) that is resistant to glyphosate inhibition. The version used in genetically modified crops was isolated from Agrobacterium strain CP4 (CP4 EPSPS) that was resistant to glyphosate. This CP4 EPSPS gene was cloned and transfected into soybeans, and in 1996, such genetically modified soybeans were made commercially available. This greatly improved the ability to control weeds in soybean fields since glyphosate could be sprayed on fields without hurting the crop. As of 2005, 87% of U.S. soybean fields were planted with glyphosate resistant varieties. [ [ IUPAC name sodium 2- [(hydroxy-oxido- phosphoryl)methylamino]acetic acid Molecular formula C 3 H 8 NO 5 P Molar mass169.07 g mol -1 Aspecto de uma vinha localizada no vale do Tejo, ao lado de um terreno destinado à plantação de tomateiros e que foi pulverizado com glifosato num dia de vento. As plantas afectadas só recuperaram após terem sido pulverizadas com uma solução aquosa de Phe + Tyr + Trp.
 that is resistant to glyphosate inhibition. The version used in genetically modified crops was isolated from Agrobacterium strain CP4 (CP4 EPSPS) that was resistant to glyphosate. This CP4 EPSPS gene was cloned and transfected into soybeans, and in 1996, such genetically modified soybeans were made commercially available. This greatly improved the ability to control weeds in soybean fields since glyphosate could be sprayed on fields without hurting the crop. As of 2005, 87% of U.S. soybean fields were planted with glyphosate resistant varieties. [ [ IUPAC name sodium 2- [(hydroxy-oxido- phosphoryl)methylamino]acetic acid Molecular formula C 3 H 8 NO 5 P Molar mass g mol -1 Aspecto de uma vinha localizada no vale do Tejo, ao lado de um terreno destinado à plantação de tomateiros e que foi pulverizado com glifosato num dia de vento. As plantas afectadas só recuperaram após terem sido pulverizadas com uma solução aquosa de Phe + Tyr + Trp..")
55
55 Mecanismo de acção do glifosato Glyphosate kills plants by inhibiting the enzyme 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS), which catalyzes the reaction of shikimate-3-phosphate (S3P) and phosphoenolpyruvate to form 5-enolpyruvyl-shikimate-3-phosphate (ESP). ESP is subsequently dephosphorylated to chorismate an essential precursor in plants for the aromatic amino acids: phenylalanine, tyrosine and tryptophan. The shikimate pathway is not present in animals, which obtain aromatic amino acids from their diet. (5-enolpyruvylshikimate- 3-phosphate synthase) Como nós não sintetizamos os aminoácidos aromáticos (isto é, não possuímos a enzima EPSPS), o glifosato não é tóxico para nós.
 Como nós não sintetizamos os aminoácidos aromáticos (isto é, não possuímos a enzima EPSPS), o glifosato não é tóxico para nós..")
56
Glyphosate and transgenic plants resistant to glyphosate Glyphosate is a broad-spectrum herbicide that poisons many plant species so it is frequently used to ‘burn down’ weeds on a field prior to the planting or emergence of crops. Before 1996, weeds were not observed to have evolved resistance to glyphosate in the field, but since then, the introduction of transgenic glyphosate tolerant crops has led to evolution of a number of resistant weeds as the result of the greatly increased use of the herbicide particularly during the post-emergent growth of the crops. Glyphosate kills plants by interfering with the synthesis of the amino acids phenylalanine, tyrosine and tryptophan. It does this by inhibiting the enzyme 5- enolpyruvylshikimate-3-phosphate synthase (EPSPS). Some microorganisms (e.g. Agrobacterium tumefaciens strain CP4) have a version of EPSPS that is resistant to glyphosate inhibition. This version was expressed in plants by genetic modification of crop plants which became resistant to glyphosate.
. Some microorganisms (e.g. Agrobacterium tumefaciens strain CP4) have a version of EPSPS that is resistant to glyphosate inhibition. This version was expressed in plants by genetic modification of crop plants which became resistant to glyphosate..")
57
Genetic basis of glyphosate tolerance The genetic basis of many of the glyphosate resistant weeds remains unknown; but those studied in detail show that there is no single genetic alteration responsible in all of the resistant weeds: - Some plants evolved partial resistance to glyphosate through changes at amino acid residue 106 of the 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) gene. Mutations change amino acid 106 from proline to either serine or threonine, results in an EPSPS weakly resistant to glyphosate. - Other plants exhibit a reduced translocation of glyphosate to meristematic tissues. - Yet other plants evolved EHSPS gene amplification. EPSPS enzyme activity from resistant and susceptible plants was equally inhibited by glyphosate. Genomes of resistant plants contained from 5-fold to more than 160-fold more copies of the EPSPS gene than did genomes of susceptible plants. In these plants, EPSPS expression was positively correlated with genomic EPSPS relative copy number. The amplified genes were not clustered on the chromosomes but distributed among all of the chromosomes. - Other plants rapidly pumped the herbicide into vacuoles preventing contact of the herbicide with the chloroplast.

58
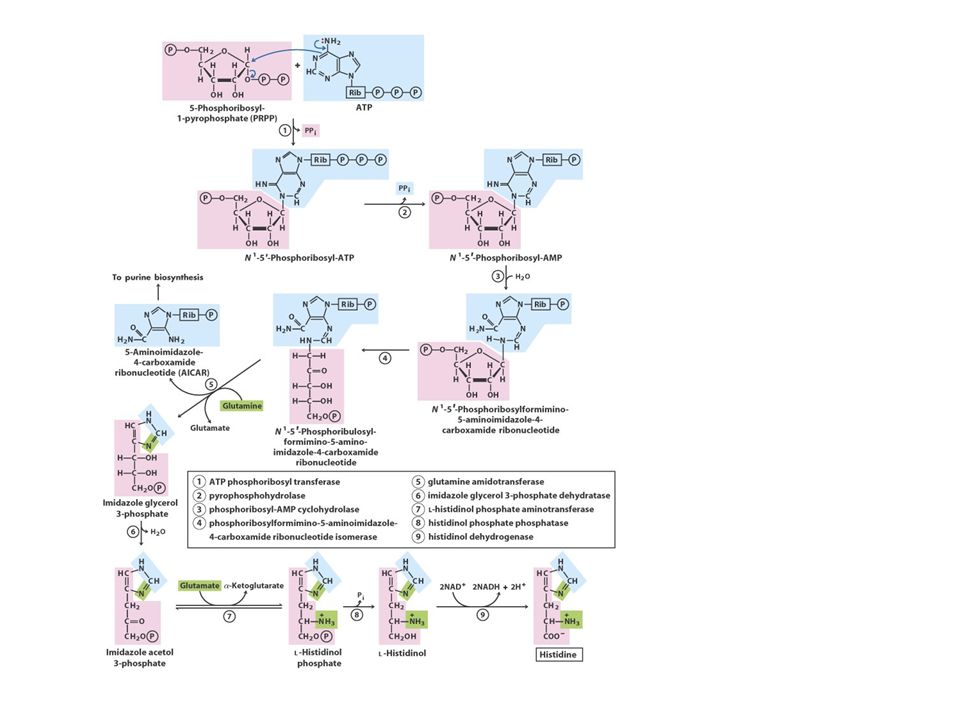
The Ribose 5-Phosphate or Histidine Family

60
Regulação da biossíntese de aminoácidos

61
Allosteric regulation of isoleucine biosynthesis Great example of this regulation (feedback inhibition)
")
62
Complex regulatory mechanisms in E.coli (aspartate as source) Note: isozymes (e.g., A 1, B 2 ) some with (e.g., C 1 ) and some without (e.g., C 2 ) allosteric regulation
 Note: isozymes (e.g., A 1, B 2 ) some with (e.g., C 1 ) and some without (e.g., C 2 ) allosteric regulation")
63
Além de oligopéptidos, polipéptidos e proteínas, os aminoácidos são precursores de uma grande variedade de compostos de grande importância biológica Synthesis of Biologically Important Compounds In addition to their role in protein synthesis, energy production and gluconeogenesis, many amino acids serve as precursors for the synthesis of other amino acids and other biologically important cornpounds. Many oligopeptides containing up to 20 residues, including hormones, antibiotics and antitumour agents, are synthesized in living organisms by mechanisms different from the usual ribosome-dependent processes of protein synthesis. The dipeptides carnosine (β-alanylhistidine) and anserine (β-alanyl-1-N-methylhistidine) are synthesized enzymatically from β-alanine and histidine, and from carnosine and S-adenosylmethionine, respectively. Glutathione (γ-glutamylcysteinylglycine) plays a variety of roles in living organisms. This tripeptide is synthesized by a two-step enzymatic pathway: (1) the formation of a peptide linkage between the γ-carboxyl group of glutamate and the amino group of cysteine, to produce γ-glutamylcysteine; (2) the condensation of this dipeptide with glycine, to form glutathione. Thus the order of the amino acids in glutathione is specifically determined by the enzymes catalysing the formation of each peptide bond. At least 90 different peptide antibiotics are produced by strains of Bacillus subtilis and B. breuis. Gramicidin S, for example, is a cyclic decapeptide composed of two identical pentapeptides ( D -phenylalanine- L -proline- L -valine- L -ornithine- L -leucine). Gramicidin S is synthesized by a multienzyme complex, gramicidin synthetase, composed of two enzyrnes, one of which, serving as a template, specifies the amino acid sequence in the antibiotic.
 and anserine (β-alanyl-1-N-methylhistidine) are synthesized enzymatically from β-alanine and histidine, and from carnosine and S-adenosylmethionine, respectively. Glutathione (γ-glutamylcysteinylglycine) plays a variety of roles in living organisms. This tripeptide is synthesized by a two-step enzymatic pathway: (1) the formation of a peptide linkage between the γ-carboxyl group of glutamate and the amino group of cysteine, to produce γ-glutamylcysteine; (2) the condensation of this dipeptide with glycine, to form glutathione. Thus the order of the amino acids in glutathione is specifically determined by the enzymes catalysing the formation of each peptide bond. At least 90 different peptide antibiotics are produced by strains of Bacillus subtilis and B. breuis. Gramicidin S, for example, is a cyclic decapeptide composed of two identical pentapeptides ( D -phenylalanine- L -proline- L -valine- L -ornithine- L -leucine). Gramicidin S is synthesized by a multienzyme complex, gramicidin synthetase, composed of two enzyrnes, one of which, serving as a template, specifies the amino acid sequence in the antibiotic..")
64
S-Adenosylmethionine (SAM), the metabolically activated form of methionine, functions as an important source of methyl and propylamino groups for a wide variety of compounds, including alkaloids, choline, creatine, adrenaline, N-methylated amino acids, nucleotides, and polyamines, as well as for phospholipids, proteins, polysaccharides, and nucleic acids, It is synthesized from methionine and ATP, in a reaction catalysed by SAM synthase. A wide variety of amines occurring in bacteria, plants and animais are derived directly or indirectly from amino acids by decarboxylation; these include ethylamine (from alanine), agmatine (from arginine), γ-aminobutyric acid (from glutamate), methylamine (from glycine), histamine (from histidine), cadaverine (from Iysine), putrescine (from ornithine), phenylethylamine (from phenylalanine), ethanolamine (from serine), tryptamine and 5- hydroxytryptamine (or serotonin; from tryptophan), and tyramine and dopamine (from tyrosine). These amines and their derivatives often play a variety of physiologically important roles. For example, γ-aminobutyric acid, phenylethylamine, tryptamine, 5-hydroxytryptamine, or serotonin, tyramine, dopamine, noradrenaline, and adrenaline are all neurologically active compounds, whereas histamine, a powerful vasodilator, is involved in allergic reactions. Tyrosine plays severaI important roles in animal metabolism as a precursor to melanins, thyroid hormones (thyroxine and triiodothyronine), and catecholamines (dopamine, noradrenaline and adrenaline). In the synthesis of melanins, tyrosinase catalyses first the hydroxylation of tyrosine to 3,4-dihydroxyphenylalanine (dopa), followed by the oxidation of dopa to phenylalanine- 3,4-quinone (dopaquinone). Dopaquinone undergoes a sequence of reactions, including polymerization, to yield both red and black melanins. Another enzyme forming dopa is tyrosine hydroxylase, which catalyses the first reaction in the sequential enzymatic pathway leading to the biosynthesis of catecholamines. Dopa is then decarboxylated to yield 3,4-dihydroxyphenylethylamine (dopamine). Dopamine is hydroxylated to norepinephrine (noradrenaline), which in turn is methylated by SAM to give epinephrine (adrenaline).
, agmatine (from arginine), γ-aminobutyric acid (from glutamate), methylamine (from glycine), histamine (from histidine), cadaverine (from Iysine), putrescine (from ornithine), phenylethylamine (from phenylalanine), ethanolamine (from serine), tryptamine and 5- hydroxytryptamine (or serotonin; from tryptophan), and tyramine and dopamine (from tyrosine). These amines and their derivatives often play a variety of physiologically important roles. For example, γ-aminobutyric acid, phenylethylamine, tryptamine, 5-hydroxytryptamine, or serotonin, tyramine, dopamine, noradrenaline, and adrenaline are all neurologically active compounds, whereas histamine, a powerful vasodilator, is involved in allergic reactions. Tyrosine plays severaI important roles in animal metabolism as a precursor to melanins, thyroid hormones (thyroxine and triiodothyronine), and catecholamines (dopamine, noradrenaline and adrenaline). In the synthesis of melanins, tyrosinase catalyses first the hydroxylation of tyrosine to 3,4-dihydroxyphenylalanine (dopa), followed by the oxidation of dopa to phenylalanine- 3,4-quinone (dopaquinone). Dopaquinone undergoes a sequence of reactions, including polymerization, to yield both red and black melanins. Another enzyme forming dopa is tyrosine hydroxylase, which catalyses the first reaction in the sequential enzymatic pathway leading to the biosynthesis of catecholamines. Dopa is then decarboxylated to yield 3,4-dihydroxyphenylethylamine (dopamine). Dopamine is hydroxylated to norepinephrine (noradrenaline), which in turn is methylated by SAM to give epinephrine (adrenaline)..")
65
Putrescine, or 1,4-diaminebutane, is synthesized by decarboxylation of ornithine, in a reaction catalysed by ornithine decarboxylase, a highly regulated enzyme. Another route to putrescine formation involves the conversion of arginine to agmatine by arginine decarboxylase, followed by cleavage of agmatine to putrescine and urea by agmatine ureohydrolase. Putrescine is an intermediate in the biosynthesis of two important polyamines, spermidine and spermine. Spermidine is synthesized enzymatically by the SAM-mediated transfer of a propylamino group to putrescine. The enzymatic transfer of an additional propylamino group from SAM to spermidine produces spermine. These polycations play multiple roles in stabilizing negatively charged intracellular components such as nucleic acids and membranes. Creatine phosphate, which serves as a source of high energy phosphate in mammalian muscle and brain, is synthesized in three steps from arginine, glycine, and methionine. There are four classes of tetrapyrrole compounds, haems, chlorophylls, phicobilins and cobalamins, all of which are synthesized from a common precursor, 6-aminolevulinic acid (ALA). In bacteria and animais, ALA is synthesized by the condensation of glycine and succinyl- CoA, with loss of carbon dioxide, in a reaction catalysed by ALA synthase. ln plants, however, ALA is formed from glutamate by a three-step pathway. An enorrnous amount of carbon in the biosphere passes through the pathway leading to lignin biosynthesis, the major constituent of woody tissue. ln the first reaction, phenylalanine ammonia Iyase catalyses the cleavage of phenylalanine to trans-cinnamic acid and NH 4 +. Cinnamic acid is a precursor for the synthesis of a huge number of plant substances, including lignin, tannins, flavonoids, pigments, many of the flavour components of spices, and various alkaloids, such as morphine and colchicine.
. In bacteria and animais, ALA is synthesized by the condensation of glycine and succinyl- CoA, with loss of carbon dioxide, in a reaction catalysed by ALA synthase. ln plants, however, ALA is formed from glutamate by a three-step pathway. An enorrnous amount of carbon in the biosphere passes through the pathway leading to lignin biosynthesis, the major constituent of woody tissue. ln the first reaction, phenylalanine ammonia Iyase catalyses the cleavage of phenylalanine to trans-cinnamic acid and NH 4 +. Cinnamic acid is a precursor for the synthesis of a huge number of plant substances, including lignin, tannins, flavonoids, pigments, many of the flavour components of spices, and various alkaloids, such as morphine and colchicine..")
66
ln addition, the synthesis of a variety of other important molecules utilizes various amino acids as precursors. Thus β-alanine is a component of CoA, asparagine is a major form of transport of organic nitrogenous compounds in plants, and aspartate is involved in purine and pyrimidine biosynthesis. Glutamate is a precursor of folic acid; glutamine contributes to the synthesis of a variety of substances, including purines, pyrimidines, ATP, cytidine triphosphate (CTP), NAD, amino sugars, and glycoproteins; cystein is a precursor of taurine, isethionic acid, CoA, vasopressin, various types of pigments, including phaeomaline and trichochromes, and other sulphur-containing compounds. Glycine also plays multiple roles, including contributions to the one-carbon pool and as a precursor of purines, glyoxylate, and various conjugates such as hippurate and glycocholate. Histidine is involved in ergothionine and homocarnosine biosynthesis, and methionine, via SAM, is the precursor of the, plant hormone ethylene, which influences plant growth and development and induces the ripening of fruits. Serine is involved in the biosynthesis of phospholipids, and tryptophan is the precursor of several important physiological substances, including NAD, NADP, and the plant hormone indole 3-acetic acid.
, NAD, amino sugars, and glycoproteins; cystein is a precursor of taurine, isethionic acid, CoA, vasopressin, various types of pigments, including phaeomaline and trichochromes, and other sulphur-containing compounds. Glycine also plays multiple roles, including contributions to the one-carbon pool and as a precursor of purines, glyoxylate, and various conjugates such as hippurate and glycocholate. Histidine is involved in ergothionine and homocarnosine biosynthesis, and methionine, via SAM, is the precursor of the, plant hormone ethylene, which influences plant growth and development and induces the ripening of fruits. Serine is involved in the biosynthesis of phospholipids, and tryptophan is the precursor of several important physiological substances, including NAD, NADP, and the plant hormone indole 3-acetic acid..")
67
Questões 1. Defina os seguintes conceitos: a) Turnover de proteínas. b) Endopeptidase. c) Ubiquitina. d) Carbamoíl-fosfato. e) Aminoácido cetogénico. f) Aminoácido essencial. g) Inibição por feedback. 2. Justifique as seguintes afirmações: a) No caso do homem, a fenilalanina é considerada um aminoácido essencial, mas não a tirosina. b) A hidrólise de proteínas é um processo exergónico: Polipéptido + (n-1) H 2 O n aminoácidos ΔG O ’ < 0 Explique, sucintamente, porque razão as células gastam energia para degradar as suas proteínas. c) O ciclo da ureia desempenha um papel essencial no metabolismo do azoto do homem. d) O glifosato, o herbicida descoberto há 40 anos e ainda o mais utilizado à escala mundial, é letal para as plantas, mas não para o homem.
 Endopeptidase. c) Ubiquitina. d) Carbamoíl-fosfato. e) Aminoácido cetogénico. f) Aminoácido essencial. g) Inibição por feedback. 2. Justifique as seguintes afirmações: a) No caso do homem, a fenilalanina é considerada um aminoácido essencial, mas não a tirosina. b) A hidrólise de proteínas é um processo exergónico: Polipéptido + (n-1) H 2 O n aminoácidos ΔG O ’ < 0 Explique, sucintamente, porque razão as células gastam energia para degradar as suas proteínas. c) O ciclo da ureia desempenha um papel essencial no metabolismo do azoto do homem. d) O glifosato, o herbicida descoberto há 40 anos e ainda o mais utilizado à escala mundial, é letal para as plantas, mas não para o homem..")
68
3. Considere a assimilação do carbono, azoto e enxofre e a biossíntese dos aminoácidos. 3.1 - Defina metabolismo do carbono, do azoto e do enxofre. 3.2 – Defina, para cada um, a forma inorgânica que é assimilada, o tipo de composto orgânico formado, a via enzimática envolvida e os organismos em que ocorre. 3.3 – Que vias metabólicas fornecem os esqueletos carbonados para a síntese dos aminoácidos? 3.4 – Identifique as seis famílias biossintéticas de aminoácidos. 3.5 – Defina, para o caso do Homem, aminoácidos não-essenciais, semi-essenciais e essenciais, dando dois exemplos de cada.

69
4 - Considere o metabolismo do azoto e dos aminoácidos. Identifique o significado das letras (nomes de enzimas, de complexos enzimáticos ou de vias metabólicas) e dos números (nomes de compostos, de famílias de compostos ou de espécies animais) na figura seguinte: Legenda:A - B - C - D - E F - G - H - I - J - K - 1 - 2 - 3 – 4 – 5 – 6 – 7 – 8 – 9 – 10 – 11 – 12 – 13 – 14 – 15 – 16 – 17 -
 e dos números (nomes de compostos, de famílias de compostos ou de espécies animais) na figura seguinte: Legenda:A - B - C - D - E F - G - H - I - J - K – 4 – 5 – 6 – 7 – 8 – 9 – 10 – 11 – 12 – 13 – 14 – 15 – 16 –")
70
FIM

Apresentações semelhantes








.>")


>")







