Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Fabrício Freitas Maria do Carmo Thiago Malardo

2
Leishmaniasis Leishmaniasis is caused by protozoan parasites belonging to the genus Leishmania. The parasites are transmitted by the bite of an insect vector, the phlebotomine sandfly. Leishmaniasis threatens about 350 million men, women and children in 88 countries around the world. As many as 12 million people are believed to be currently infected, with about 1–2 million estimated new cases occurring every year.

3
Parasite's life cycle The promastigote form of L. donovani is transmitted into the skin by female phlebotomine sandflies. Once transmitted, the parasites are internalized by dendritic cells and macrophages in the dermis where they lose their flagella, transforming into the amastigote form. The amastigotes multiply, destroy the host cell and infect other phagocytic cells. The amastigotes disseminate through the lymphatic and vascular systems, eventually infiltrating the bone marrow, liver and spleen.

4
Forms of leishmaniasis
Cutaneous leishmaniasis - is the most common form. It usually produces ulcers on the exposed parts of the body, such as the face, arms and legs; Diffuse (disseminated) cutaneous leishmaniasis - is characterized by the presence of a large (>10) number of lesions at several anatomic sites (head, limbs, and trunk); Mucocutaneous leishmaniasis - the lesions can lead to partial or total destruction of the mucous membranes of the nose, mouth and throat cavities and surrounding tissues; Visceral leishmaniasis - is the most severe form, in which vital organs of the body are affected.
 cutaneous leishmaniasis - is characterized by the presence of a large (>10) number of lesions at several anatomic sites (head, limbs, and trunk); Mucocutaneous leishmaniasis - the lesions can lead to partial or total destruction of the mucous membranes of the nose, mouth and throat cavities and surrounding tissues; Visceral leishmaniasis - is the most severe form, in which vital organs of the body are affected.")
5
Global distribution of leishmaniasis
BMJ VOLUME FEBRUARY 2003

6
Geographical distribution
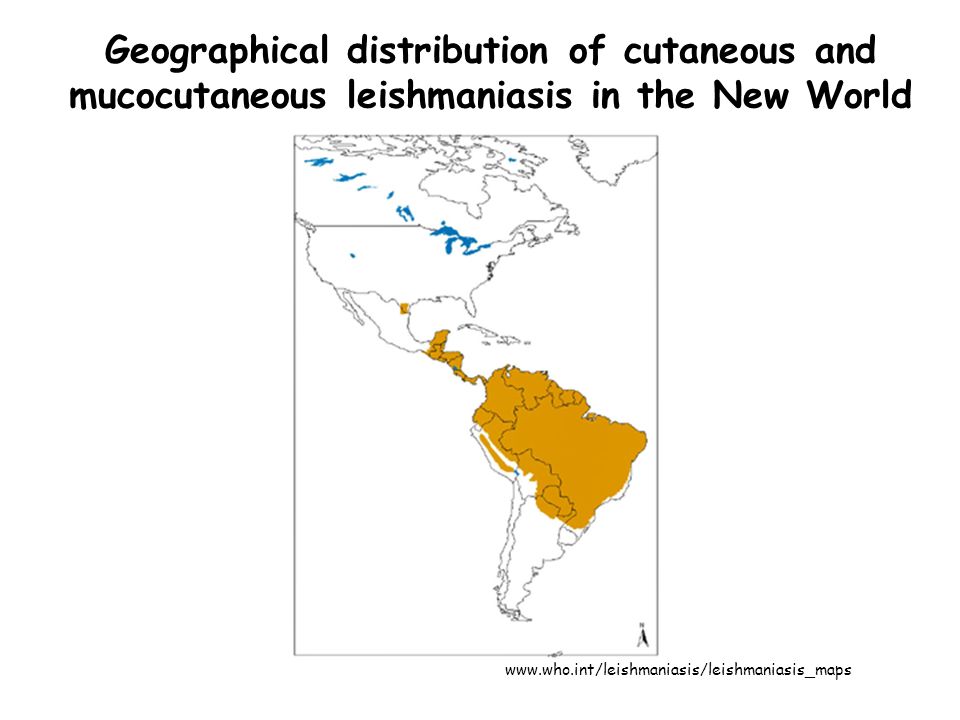
90% of cutaneous leishmaniasis cases occur in Afghanistan, Brazil, Iran, Peru, Saudi Arabia and Syria; 90% of mucocutaneous leishmaniasis occurs in Bolivia, Brazil and Peru; 90% of all visceral leishmaniasis cases occur in Bangladesh, Brazil, India, Nepal and Sudan.

7
Geographical distribution of cutaneous and mucocutaneous leishmaniasis in the New World
8
The main Leishmania species

9
Treatment regimens

10
Immune response against Leishmania

11
TLR and MyD88 pathway in Leishmania infection

12
TLR signaling pathway K. Takeda, S. Akira / Seminars in Immunology 16 (2004) 3–9
 3–9")
13
Events that lead to susceptibility
Late events in susceptible mice. The evolution of susceptibility to Leishmania major, as occurs after sub-cutaneous (s.c.) inoculation of a relatively high number of parasites into BALB/c mice, or intravenous (i.v.) or intranasal (i.n.) inoculation of a high number of parasites into C57BL/6 mice, is due to the failure of an interleukin-12 (IL-12)-dependent redirection of the early T helper 2 (TH2) response, resulting in the clonal expansion and dominance of CD4+CD45RBlow TH2 cells. The TH1-response defect might be due to several mechanisms, including: a | dissemination of parasites to the viscera, where TH2-priming conditions are maintained. The class of immune response in these tissues might be biased by the presence of lineage-specific type-2 dendritic cells (DC2s) that express the appropriate co-stimulatory molecules and cytokines required to drive TH2-cell development, or by the cytokine milieu that conditions DCs to promote TH2-cell development; and/or b | unstable expression of IL-12 receptor β2-chain (IL-12Rβ2) on activated CD4+ T cells. Type-2 cytokines (c) contribute to susceptibility by downregulating TH1-cell differentiation and by conditioning infected macrophages to become unresponsive to activation signals required for nitric oxide (NO)-dependent killing. Sustained neutrophil recruitment to the inflammatory site (d) might be inhibitory to TH1-cell development, owing, for example, to the local secretion of transforming growth factor-β (TGF-β) or IL-10. IL-10 produced by macrophages as a consequence of Fc-receptor (FcR) ligation by parasite-specific antibodies (e) might contribute to the pool of deactivating cytokines in infected tissues. A minor population of interferon-γ (IFN-γ)-producing CD4+CD45RBhi effector cells (f) remain active even during the progressive stages of disease. IgG, immunoglobulin G; iNOS, inducible nitric oxide synthase.
 inoculation of a relatively high number of parasites into BALB/c mice, or intravenous (i.v.) or intranasal (i.n.) inoculation of a high. number of parasites into C57BL/6 mice, is due to the failure of an interleukin-12 (IL-12)-dependent redirection of the early T helper 2. (TH2) response, resulting in the clonal expansion and dominance of CD4+CD45RBlow TH2 cells. The TH1-response defect might be. due to several mechanisms, including: a | dissemination of parasites to the viscera, where TH2-priming conditions are maintained. The class of immune response in these tissues might be biased by the presence of lineage-specific type-2 dendritic cells (DC2s) that. express the appropriate co-stimulatory molecules and cytokines required to drive TH2-cell development, or by the cytokine milieu. that conditions DCs to promote TH2-cell development; and/or b | unstable expression of IL-12 receptor β2-chain (IL-12Rβ2) on. activated CD4+ T cells. Type-2 cytokines (c) contribute to susceptibility by downregulating TH1-cell differentiation and by conditioning. infected macrophages to become unresponsive to activation signals required for nitric oxide (NO)-dependent killing. Sustained. neutrophil recruitment to the inflammatory site (d) might be inhibitory to TH1-cell development, owing, for example, to the local. secretion of transforming growth factor-β (TGF-β) or IL-10. IL-10 produced by macrophages as a consequence of Fc-receptor (FcR) ligation by parasite-specific antibodies (e) might contribute to the pool of deactivating cytokines in infected tissues. A minor. population of interferon-γ (IFN-γ)-producing CD4+CD45RBhi effector cells (f) remain active even during the progressive stages of. disease. IgG, immunoglobulin G; iNOS, inducible nitric oxide synthase.")
14
Events that lead to resistance
Late events in resistant mice. The evolution of resistance to Leishmania major, as occurs after sub-cutaneous (s.c.) or intradermal (i.d.) inoculation of a high or low number of parasites in C57BL/6 mice, or a low number of parasites in BALB/c mice, also involves the early production of interleukin-4 (IL-4) and other type-2 cytokines by CD4+ T cells. However, the parasites remain restricted to the site of infection and to the local draining lymph nodes, where, in response to accumulating amastigotes and endogenous agonists such as CD40L and interferon-γ (IFN-γ), dendritic cells (DCs) are activated to upregulate expression of CD40 and other co-stimulatory molecules and to produce IL-12 (a). These local priming conditions generate a predominant T helper 1 (TH1) response characterized by CD4+CD25−CD45RBhi effector cells that produce high levels of IFN-γ and tumour-necrosis factor (TNF) to upregulate the expression of inducible nitric oxide synthase (iNOS) and activate infected macrophages for intracellular killing (b). CD8+ T cells cooperate to control infection by their production of IFN-γ and by their lytic activities, which might be mediated by a Fas–Fas ligand (FasL)-dependent pathway (c). After healing, low numbers of amastigotes persist at the site of infection in macrophages, DCs and fibroblasts, owing (devido) to the production of IL-10 by CD4+CD25+ regulatory T cells (d).
 or intradermal (i.d.) inoculation of a high or low number of parasites in C57BL/6 mice, or a low number of parasites in BALB/c mice, also involves the early production of interleukin-4 (IL-4) and other type-2 cytokines by CD4+ T cells. However, the parasites remain. restricted to the site of infection and to the local draining lymph nodes, where, in response to accumulating amastigotes and. endogenous agonists such as CD40L and interferon-γ (IFN-γ), dendritic cells (DCs) are activated to upregulate expression of CD40. and other co-stimulatory molecules and to produce IL-12 (a). These local priming conditions generate a predominant T helper 1. (TH1) response characterized by CD4+CD25−CD45RBhi effector cells that produce high levels of IFN-γ and tumour-necrosis factor. (TNF) to upregulate the expression of inducible nitric oxide synthase (iNOS) and activate infected macrophages for intracellular killing (b). CD8+ T cells cooperate to control infection by their production of IFN-γ and by their lytic activities, which might be mediated by a. Fas–Fas ligand (FasL)-dependent pathway (c). After healing, low numbers of amastigotes persist at the site of infection in. macrophages, DCs and fibroblasts, owing (devido) to the production of IL-10 by CD4+CD25+ regulatory T cells (d).")
15
Resistance and susceptibility in leishmaniasis
Model of TH1-/TH2-cell development. For both T helper 1 (TH1)- and TH2-cell differentiation, antigens are presented to naive CD4+ T cells by dendritic cells (DCs). The interaction of co-stimulatory molecules with their respective ligands (CD40–CD40L, OX40–OX40L and/or CD80–CTLA4/CD28), together with the local cytokine environment, promotes the differentiation of naive T cells into interferon-γ (IFN-γ)-secreting TH1 cells or interleukin-4 (IL-4)-secreting TH2 cells. It has been proposed also that distinct subsets of dendritic cells, known as DC1 and DC2, might exist, which, in turn, direct TH1 and TH2 differentiation pathways, respectively. a | In TH1- cell development, certain pathogens or pathogen-associated molecular patterns (PAMPs) trigger antigen-presenting cells, through Toll-like receptors (TLRs), to secrete IL-12, which promotes the differentiation of naive T cells into IFN-γ-secreting TH1 cells. Signal transducer and activator of transcription 4 (STAT4) and STAT1 are activated by IL-12 and IFN-γ, respectively. T-bet, a T-box transcription factor is TH1-lineage specific. b | In TH2-cell development, the inability of antigen to activate DCs to produce IL-12 results in a default pathway of naive T-cell differentiation into IL-4-secreting TH2 cells. In addition, antigen and/or specific tissue environments might activate DCs to produce IL-4 or IL-10, which will instruct TH2-cell development. STAT6 is activated specifically by IL-4-receptor binding. The TH2-lineage-specific transcription factor GATA3 binds to consensus GATA-binding sites (AGATAG). The factors c-Maf and NFATc (nuclear factor of activated T cells) have been associated with TH2 differentiation also. Bcl-6, ROG (repressor of GATA) and FOG1 (friend of GATA1) negatively regulate TH2 differentiation by repressing the activity of STAT6 and GATA3. Suppressor of cytokine signalling (SOCS)-family members inhibit TH1 and TH2 responses by blocking STAT activity. CTLA4, cytotoxic T-lymphocyte antigen 4; TCR, T-cell receptor; TGF-β, transforming growth factor-β; TNF, tumour-necrosis factor. Suppressor of cytokine signalling (SOCS) NATURE REVIEWS / IMMUNOLOGY / VOLUME 2 / NOVEMBER 2002
- and TH2-cell differentiation, antigens are presented to. naive CD4+ T cells by dendritic cells (DCs). The interaction of co-stimulatory molecules with their respective ligands (CD40–CD40L, OX40–OX40L and/or CD80–CTLA4/CD28), together with the local cytokine environment, promotes the differentiation of naive T cells. into interferon-γ (IFN-γ)-secreting TH1 cells or interleukin-4 (IL-4)-secreting TH2 cells. It has been proposed also that distinct subsets of. dendritic cells, known as DC1 and DC2, might exist, which, in turn, direct TH1 and TH2 differentiation pathways, respectively. a | In TH1- cell development, certain pathogens or pathogen-associated molecular patterns (PAMPs) trigger antigen-presenting cells, through. Toll-like receptors (TLRs), to secrete IL-12, which promotes the differentiation of naive T cells into IFN-γ-secreting TH1 cells. Signal. transducer and activator of transcription 4 (STAT4) and STAT1 are activated by IL-12 and IFN-γ, respectively. T-bet, a T-box. transcription factor is TH1-lineage specific. b | In TH2-cell development, the inability of antigen to activate DCs to produce IL-12 results. in a default pathway of naive T-cell differentiation into IL-4-secreting TH2 cells. In addition, antigen and/or specific tissue environments. might activate DCs to produce IL-4 or IL-10, which will instruct TH2-cell development. STAT6 is activated specifically by IL-4-receptor. binding. The TH2-lineage-specific transcription factor GATA3 binds to consensus GATA-binding sites (AGATAG). The factors c-Maf. and NFATc (nuclear factor of activated T cells) have been associated with TH2 differentiation also. Bcl-6, ROG (repressor of GATA) and. FOG1 (friend of GATA1) negatively regulate TH2 differentiation by repressing the activity of STAT6 and GATA3. Suppressor of cytokine. signalling (SOCS)-family members inhibit TH1 and TH2 responses by blocking STAT activity. CTLA4, cytotoxic T-lymphocyte antigen 4; TCR, T-cell receptor; TGF-β, transforming growth factor-β; TNF, tumour-necrosis factor. Suppressor of cytokine signalling (SOCS) NATURE REVIEWS / IMMUNOLOGY / VOLUME 2 / NOVEMBER")
16
Thiago Malardo

17
Why some strains of Leishmania cause more severe injuries than others

18
Parasitized parasite Entamoeba histolytica (1960);
Giardia lamblia (1986); Trichomonas vaginalis (1986); Most recently in Leishmania (1988).
; Trichomonas vaginalis (1986); Most recently in Leishmania (1988).")
20
Leishmania infected by virus
Virus - Double-stranded RNA (dsRNA); Viruses of Leishmania have been identified in 13 strains of L. braziliensis or L. guyanensis; Are designated Leishmania RNA virus 1 (LRV1; Totiviridae).
; Viruses of Leishmania have been identified in 13 strains of L. braziliensis or L. guyanensis; Are designated Leishmania RNA virus 1 (LRV1; Totiviridae).")
21
Levels of RNA virus within the parasite affect the host`s immune response

22
Cutaneous leishmaniasis

23
Mucocutaneous leishmaniasis

24
Fabrício Freitas

25
Objetivo Analisar o papel do TLR 2 e MyD88 na ativação de células dentríticas durante a infecção por Leishmania braziliensis

26
A produção de citocinas e ativação das MyD88-/- DC estão prejudicadas após a infecção por Leishmania braziliensis ? BMDC WT MyD88-/- Citometria de fluxo ELISA Infecção 8:1 24 horas L. braziliensis (Lb) L. amazonensis (La)
 L. amazonensis (La)")
27
A produção de citocinas e ativação das MyD88-/- DC estão prejudicadas após a infecção por Leishmania braziliensis ? Produção de IL-12p40 e ativação da DC foi prejudicada na ausência de MyD88.

28
Camundongos MyD88-/- são mais susceptíveis à infecção por Leishmania braziliensis ?
WT ou MyD88-/- 2x106 Pata posterior L. braziliensis semanalmente Lesão Camundongos MyD88-/- são mais susceptíveis a infecção por L. braziliensis

29
Qual o mecanismo do MyD88 na susceptibilidade à infecção por Leishmania braziliensis ?
WT ou MyD88-/- 2x106 Pata posterior L. braziliensis 4 e 8 semanas Linfonodo drenante 1x106 cels. 6 h Citometria CD4+ (IFN-γ, IL-17 e IL-10) O mecanismo do MyD88 é mediar a produção de céls. TCD4+ produtoras de IFN-γ e IL-17, sendo necessário para uma efetiva resposta imune contra L.braziliensis.
 O mecanismo do MyD88 é mediar a produção de céls. TCD4+ produtoras de IFN-γ e IL-17, sendo necessário para uma efetiva resposta imune contra L.braziliensis.")
30
Toll 2 molécula chave na intereção hospedeiro-Leishmania.

31
Toll 2 está envolvido no reconhecimento da Leishmania braziliensis ?
BMDC WT Toll 2-/- Citometria de fluxo ELISA Infecção 8:1 24 horas L. braziliensis (Lb) L. amazonensis (La)
 L. amazonensis (La)")
32
Toll 2 está envolvido no reconhecimento da Leishmania braziliensis ?
Na ausência de Toll2, DC são mais ativadas e produzem maiores quantidades de IFN-γ.

33
O aumento da ativação de toll 2-/- DC está associada com a expressão de SOCS após a infecção por Leishmania braziliensis ? BMDC WT Toll 2-/- Infecção 10:1 4 horas Real-Time PCR L. braziliensis (Lb) L. amazonensis (La)
 L. amazonensis (La)")
34
O aumento da ativação de toll 2-/- DC está associada com a expressão de SOCS após a infecção por Leishmania braziliensis ? Na ausência de Toll2, DC expressaram maiores níveis de SOCS 1 e 3. Regulação positiva independente de toll 2.

35
Toll 2-/- DC infectadas com Leishmania braziliensis podem ativar células T CD4+produtoras de IFN-γ ?
BMDC WT Toll 2-/- CD4+ baço 2x106 Infecção 8:1 24 horas Co-cultura 4 dias Proliferação ELISA L. braziliensis (Lb) L. amazonensis (La)
 L. amazonensis (La)")
36
Toll 2-/- DC infectadas com Leishmania braziliensis podem ativar células T CD4+produtoras de IFN-γ ?
A deficiência de toll 2 promove ativação da céls. T CD4+e a produção de IFN-γ.

37
Camundongos Toll 2-/- são mais resistentes à infecção por Leishmania braziliensis ?
WT ou Toll 2-/- 2x106 Pata posterior L. braziliensis semanalmente Lesão Camundongos Toll 2-/- são mais susceptíveis a infecção por L. braziliensis.

38
Como estaria a carga parasitária em camundongos Toll 2-/- infectados com Leishmania braziliensis ?
WT ou MyD88-/- 2x106 Pata posterior L. braziliensis 4 e 8 semanas Carga parasitária No grupo WT e K.O, houve semelhante carga parasitária na quarta e oitava semana.

39
A proteção em animais toll 2-/- estaria associada com o aumento de céls. T CD4+ produtoras de IFN-γ?
WT ou MyD88-/- 2x106 Pata posterior L. braziliensis 4 e 8 semanas Linfonodo drenante 1x106 cels. 6 h Citometria CD4+ (IFN-γ, IL-17 e IL-10)
")
40
A proteção em animais toll 2-/- estaria associada com o aumento de céls. T CD4+ produtoras de IFN-γ?
A proteção está associada com um aumento da população de céls. T CD4+ produtoras de IFN-γ

41
A proteção em animais toll 2-/- estaria associada com a alta produção de IFN-γ pelas céls. T CD4+ ?
WT ou MyD88-/- 2x106 Pata posterior L. braziliensis 4 e 8 semanas Linfonodo drenante 1x106 cels. SLA 3 dias Elisa (IFN-γ e IL-10)
")
42
A proteção em animais toll 2-/- estaria associada com a alta produção de IFN-γ pelas céls. T CD4+ ?
A proteção está associada com a alta produção de IFN-γ pelas céls. T CD4+

43
Conclusão PAPEL REGULADOR
2 Elevada ativação e produção de IL-12 e aumento na população de células T CD4+ produtoras de IFN-γ. PAPEL REGULADOR A falta do MyD88 prejudica a imunidade protetora no combate à infecção por Leishmania braziliensis.

44
Objetivo: Verificar a modulação da resposta imune por diferentes linhagens de Leishmania guyanensis e o envolvimento do LRV1 nesta modulação.

45
As linhagens L.g.M+ e L.g.M- modulam diferencialmente a resposta dos MΦ hospedeiro?
BMMΦ (BALB/c) 6h DNA microarrays – expressão de genes (RI)
 6h. DNA microarrays – expressão de genes (RI)")
46
As linhagens L.g.M+ e L.g.M- modulam diferencialmente a resposta dos MΦ hospedeiro?
L.Major LV39 BMMΦ (BALB/c ou C57BL/6) 6h ELISA
 6h. ELISA.")
47
Linhagens isoladas CL e CLM também modulam diferencialmente a resposta dos MΦ hospedeiro?
(BALB/c ou C57BL/6) 6h L.g. h-CL (Lg1881) L.g. h-MCL (Lg1398) ELISA
 6h. L.g. h-CL (Lg1881) L.g. h-MCL (Lg1398) ELISA.")
48
L.g.M+ M5313 L.g. h-MCL (Lg1398) L.g.M- L.g. h-CL (Lg1881)
Metastáticas CXCL10, CCL5, TNF-α e IL-6 L.g.M- L.g. h-CL (Lg1881) Não- metastáticas CXCL10, CCL5, TNF-α e IL-6 Os elevados níveis de citocinas e quimiocinas, depois da infecção, estão associados com as linhagens de parasitas metastáticos.
 Não- metastáticas. CXCL10, CCL5, TNF-α e IL-6. Os elevados níveis de citocinas e quimiocinas, depois da infecção, estão associados com as linhagens de parasitas metastáticos.")
49
Cloroquina ou Citocalasina D
A entrada do parasita e a formação de fagolisossomo são necessários para o desenvolvimento desta resposta? L.g.M+ L.g.M- M5313 BMMΦ (C57BL/6) Cloroquina ou Citocalasina D 6h ELISA
 Cloroquina ou Citocalasina D. 6h. ELISA.")
50
Existe envolvimento dos TLRs endossomais (TLR3, 7 e 9) nesta resposta?
BMMΦ C57BL/6 TLR3-/- TLR7-/- TLR9-/- L.g.M+ L.g.M- M5313 6h ELISA

51
TLR3 é o receptor responsável pelo reconhecimento do parasita?
BMMΦ C57BL/6 TLR3-/-TRIF-/- MyD88-/- L.g.M+ L.g.M- M5313 6h ELISA (IFN-β) A via dependente de TRL3-TRIF foi essencial para o aumento da expressão dos mediadores proinflamatórios
 A via dependente de TRL3-TRIF foi essencial para o aumento da expressão dos mediadores proinflamatórios.")
52
Os MΦ reconhecem dsRNA das linhagens metastaticas?
Ácidos nucléicos L.m.LV M5313 L.g.M+ L.g.M- ssRNAse ou DNAse/ssRNAse BMMΦ BALB/c 6h ELISA

53
L.g.M5313 (M+) L. guyanensis L. braziliensis dsRNA Leishmania RNA virus 1 (LRV1) Isolados de MCL humana
 Isolados de MCL humana.")
54
O LRV1 esta presente nas linhagens metastaticas?
Ácidos nucléicos L.g.M- L.g.M+ M5313 L.g. h-CL (Lg1881) L.g. h-MCL (Lg1398) L.m.LV39 DNAse ou não ssRNAse A RNA total Real Time PCR B
 L.g. h-MCL (Lg1398) L.m.LV39. DNAse ou não. ssRNAse. A. RNA total. Real Time PCR. B.")
55
O LRV1 é o responsável pelo aumento de citocinas e quimiocinas na infecção com linhagens metastáticas? BMMΦ C57BL/6 TLR3-/- dsRNA LRV1 Real Time PCR

56
Estes resultados são similares para linhagens isogênicas?
BMMΦ C57BL/6 TLR3-/- L.g.M4147-LRVhigh L.g.M4147-LRVneg Real Time PCR

57
TLR3 e LRV1 desenvolvem o mesmo papel na leishmaniose in vivo?
Camundongos C57BL/6 TLR3-/ TLR7-/- Footpad swelling A L.g.M+ L.g.M- Carga parasitária (qRT-PCR) B
 B.")
58
Conclusão

Apresentações semelhantes











>")

>")






: Descoberta, Estrutura, Organização Genômica e Regulação da Expressão das Moléculas do MHC QUESTÕES.>")