Carregar apresentação
A apresentação está carregando. Por favor, espere

1
A mecânica da divisão celular: Mitose
UNIVERSIDADE ESTADUAL DE CAMPINAS Programa de Pós- Graduação em Biologia Celular e Estrutural A mecânica da divisão celular: Mitose Cristiano Pedrozo Vieira¹; Dra. Shirlei Maria Recco-Pimentel² ¹ Aluno do Programa de Pós-Graduação em Biologia Celular e Estrutural. Instituto de Biologia/Unicamp. Seminário apresentado na disciplina de Biologia Celular, N716. ² Orientadora do seminário. Disciplina de Biologia Celular, N716. Instituto de Biologia/Unicamp

2
Introdução

3
Introdução G1 – intensa síntese de componentes necessários para a divisão ocorrer. A célula dobra de volume. Intensa transcrição, tradução Multiplicação de organelas Aumento da membrana plasmática. aumento do citoesqueleto. S Replicacao do DNA, proteinas histonicas.

4
Introdução Pelo tempo: fibroblastos.

5
Todas as células se dividem ?
Introdução Células que nunca dividem neurônios, muscular, hemácias Células que raramente dividem Todas as células se dividem ? hepatócitos, pâncreas Células que dividem muito espermatogônias, células tronco em geral (dos epitélios, da medula óssea etc.)
")
6
subunidades da condensina Lamina nuclear que sustenta EN
M-CDK * desencadeia uma cascata de fosforilação das proteínas que iniciam a mitose. * Essas fosforilações são responsáveis por mudanças morfológicas que ocorrem durante a mitose. Fosforilação subunidades da condensina Cromossomos se condensam A M-CDK ativada fosforila algumas subunidades da condensina, desencadeando a associação do complexo condensina no DNA, portanto a progressiva condensação do cromossomo. As condesinas podem utilizar energia da hidrólise do ATP para promover o enrolamento do DNA. - Na súbita transição da metafase-anáfase inicia a desfosforilação de muitas proteínas que estavam fosforiladas na prófase. Após o desligamento da M-Cdk a fosfatase passa agir sem oposição. Lamina nuclear que sustenta EN EN se rompe

7
Introdução Células de pulmão de salamandra
Micrografia de cultura de células de pulmão de salamandra: Microtubulos ( verde) visualizados por imunofluorescência. Cromátide é fixada com um corante fluorescente azul. - Comportamento de microtúbulos e da cromatina. Células de pulmão de salamandra
 visualizados por imunofluorescência. Cromátide é fixada com um corante fluorescente azul. - Comportamento de microtúbulos e da cromatina. Células de pulmão de salamandra.")
8
Duas maquinarias citoesqueléticas na fase M
Introdução Duas maquinarias citoesqueléticas na fase M Duas maquinárias citoesqueléticas operam na fase M. O fuso mitotico agrupa-se e segrega os cromossomos. O anel contrátil agrupa-se para dividir a célula em duas. O ciclo do centrossomo. As células vegetais utilizam um mecanismo muito diferente para dividir o citosol.

9
Ciclo do centrossomo Camundogo (929)
A duplicação dos centriolos é um mecanismo desconhecido. A replicacao dos centriolos consiste de um par de centriolos associadaos a matriz [ verde] Em g1, os dois centriolos do par se separam poucos micrometros. Na fase S: o centriolo filho começa a crescer próximo a base de cada centriolo-mae, em um ângulo de 90 graus com este A elongacao do centriolo – filho é normalmente finalizada em G2. Os dois pares de centriolos permanevem juntos em um único complemento centrossomal até o início da fase M, quando o complexo se divide em dois, e as duas metades começam a se separar. Cada centrossomo agora, forma o seu próprio arranjo radial de microtubulos, denominado áster. Camundogo (929)
")
10
A A – micrografia eletronica da fase S de uma célula mamifera em cultura, apresentando um centrossomo duplicado. Cada centrossomo contém um par de centriolos, embora os centriolos tenham se duplicado, eles permanecem juntos em único complexo, como mostrado em B [ bege] B – um centriolo de cada par de centriolos é cortado, em uma secção transversal, enquanto o outro é cortado em uma secção logitudinal, indicando que os dois membros de cada par estão alinhados em ângulo reto. As duas metades do centrossomos replicado, consistindo cada uma de par de centriolos rodeados por pela matriz,. Irão se separar e migrarão para iniciar a formação dos dois pólos do fuso, quando a célula entrar na fase M C - par de centriolo que foi isolado de uma célula. Os dois centriolos foram parcialmente separados, durante o procedimento de isolamento, mas permanecem amarrados juntos por fibras finas, as quais mantém o par de centriolos unidos até chegar o momento da separação. Ambos os centriolos são cortados longitudinalmente e agora pode ser vistos que os dois apresentam estruturas diferentes: o centriolo-mae é maior e mais complexo que o centriolo-filha e, como mostrado ANTES, somente o centriolo mae está associado a matriz que forma os microtubulos. Cada centriolo-filho irá amadurecer durante o proximo ciclo celular quando irá se replicar C B

11
Coesina e condensina O complexo depositado na fase G1 e a coesão é estabelecida concomitante a replicação DNA. A ligação física entre as cromátides irmãs é mantida em G2. Associação das cromátides irmãs depende das COESINAS - complexo protéico com 4 subunidades- .

12
Condesina é inativada no final da divisao celular
Ação de fosfatases . - Se a coesao da cromatide ñ está estabilizada corretamente na fase S, a completa condensacao não pode ocorrer na fase M, e os cromossomos serão irregularmente segregados na anáfase.

13
Modelos da interação da Coesina
- Recrutamento da coesina a regiões distintas em centromeros, necessitam de proteinas acessórias. O complexo depositado na fase G1 e a coesão é estabelecida concomitante a replicação DNA. A ligação física entre as cromátides irmãs é mantida em G2. Populações diferentes da coesina existem em diferentes regiões dos cromossomos. A distribuição fiel dos cromossomos necessitam que as cromatides irmas estão seguradamente juntas da fase S ao mesmo tempo na mitose. A maioria das coesinas se dissocia da cromatina durante a prófase, em processo regulado por quinases: Aurora B e polo quinase. Considerando uma pequena população na heterocromatina pericentrica que permanace até anáfase. Losada., 2007

14
Modelos da interação da Coesina
Presença de proteinas da familia shugoshin [ sob investigacoes] essencial para proteger a coesina centromérica da dissociacão na profase TRF1 protege o complexo telomérico Proteinas modificadas no centromero que promovem interacao mais forte Losada., 2007

15
Condensina Cromossomo bem condensado! A condensação de cromossomos na mitose depende das CONDENSINAS, complexo protéico com 5 subunidades Eixo fixado com um anticorpo fluorescente contra uma proteina do complexo condensina DNA

16
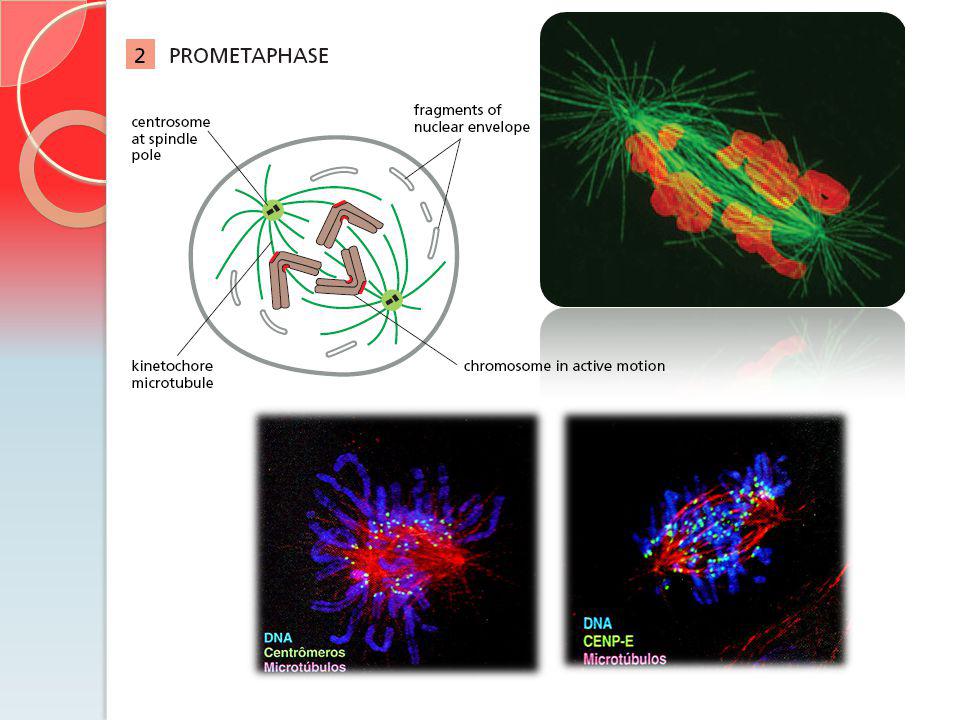
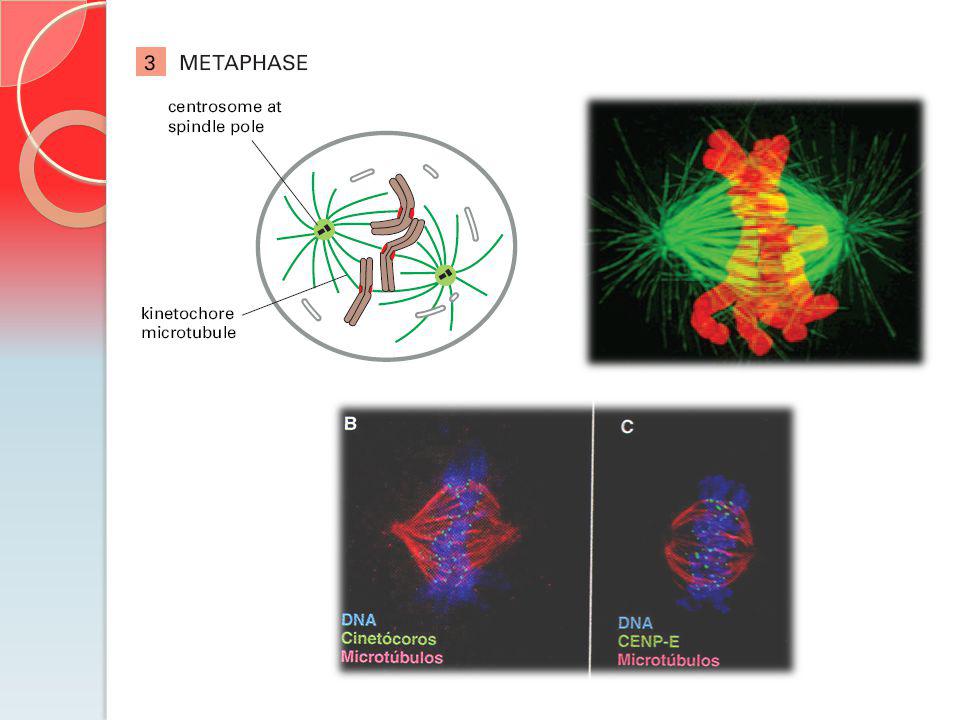
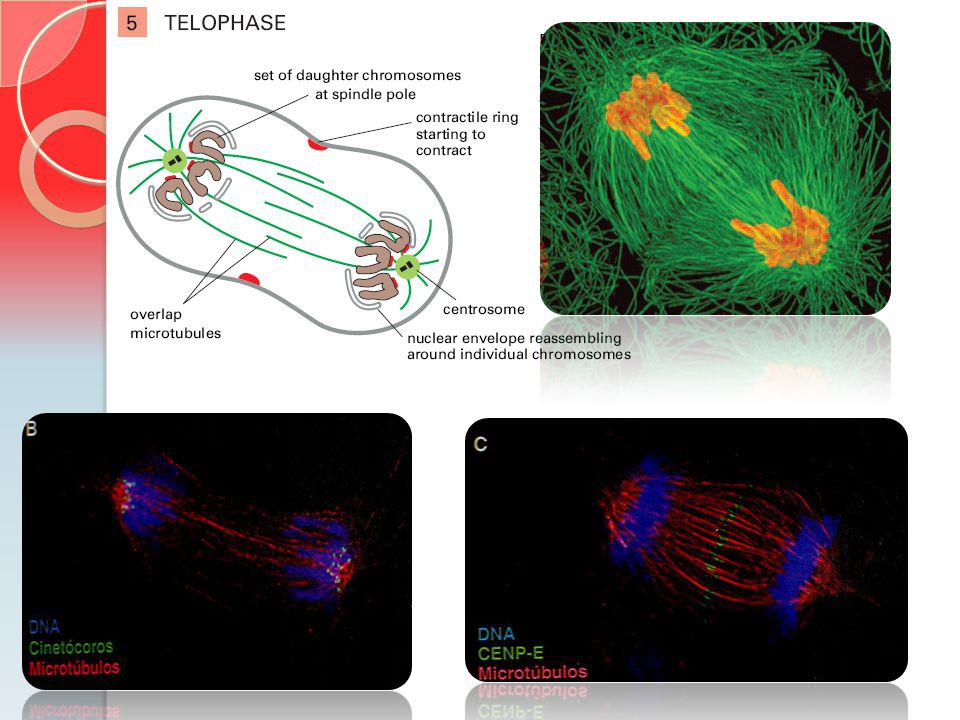
Estágios da Mitose CENP-E = Cinesina que está envolvida no monitoramento da ligação dos cinetócoros do fuso. CENP-E = grande cinesina que está envolvida no monitoramento da ligação dos cinetócoros do fuso Cs = montagem do fuso central

18
Cinetocoro 1 a 40 MT cinetocoro

19
Fluxo em direção ao pólo: Cinetocoros e Microtúbulos
Procura e captura. Microtúbulos nucleados a partir de cada centrossomo que está se separando crescem em direção ao cromossomo. Rede de tubulinas adicionadas na extremidade +, equilibrando a perda nas extremidades -, próximo ao pólo do fuso. Fluxo em direção ao pólo: Cinetocoros e Microtúbulos

20
Alga.

21
Duas forças opostas Um modelo de como duas forcas opostas podem cooperar para mover os cromossomos para a placa metafasica.

22
Microtúbulos

23
Equilíbrio entre MAP e catastrofinas
Dinâmica de Microtúbulos Extrato de ovos de Xenopus incubados com centrossomos Evidência experimental de que o equilibrio entre as catastrofinas e as Maps influencia a frequência das catastrófes e dos alongamentos dos microtúbulos. Extratos de ovos de xenopus na interfase mitotica foram incubados com centrossomos e o comportamento dos microtubulos individuais formados a partir do centrossomo foram monitorados por microscopia de videofluorescencia. A taxa de catastrofes é maior em extratos mitoticos do que nas interfase. A depleção de uma MAP especifica ( XMAP215) do extrato mitótico aumenta extraordinariamente a taxa de catastrófe, indicando que esta MAP, normalmente, inibe as catastrófes no extrato mitótico. A adição de anticorpos que bloqueiam a função de uma catastrofina especifica, reduz a taxa de catastrófes em extratos mitóticos Depende o equílibrio entre a MAP e as catastrofinas. A micrografia de fluorescencia dos ásteres formados em diferentes condições experimentais é apresentada, o aumento na taxa de catastrofes encurta os microtubulos. Equilíbrio entre MAP e catastrofinas
 do extrato mitótico aumenta extraordinariamente a taxa de catastrófe, indicando que esta MAP, normalmente, inibe as catastrófes no extrato mitótico. A adição de anticorpos que bloqueiam a função de uma catastrofina especifica, reduz a taxa de catastrófes em extratos mitóticos. Depende o equílibrio entre a MAP e as catastrofinas. A micrografia de fluorescencia dos ásteres formados em diferentes condições experimentais é apresentada, o aumento na taxa de catastrofes encurta os microtubulos. Equilíbrio entre MAP e catastrofinas.")
24
Influência das proteínas motoras opostas no alongamento do fuso das leveduras de brotamento
Equilíbrio entre proteínas motoras opostas determina o tamanho do fuso nestas células. A – micrografia de contraste de interferência diferencial. CID O fuso está destacado em verde, e a posição dos pólos do fuso está indicada por setas vermelhas. O envelope nuclear não se rompe durante a mitose de leveduras, e o fuso se forma dentro do núcleo. B-D o fuso mitotico foi fixado com anticorpos fluorescentes antitubulina. B – celula de levedura normal. C – a surperexpressão da proteina Kar3p, direcionada na extremidade menos, conduz a um encurtamento anormal do fuso. D- as surprexpressao da proteina motora Cin8p, direcionada na extremidade mais, conduz a um alongamento anormal do fuso. Isso demonstra que um equilibrio entre proteinas motoras opostas determina o tamanho do fuso nestas células. Extremidade menos Extremidade mais

26
MT do cinetocoro estão estáveis.
Microtúbulos na metáfase não mudam o seu comprimento: quantidade de tubulina que entra na extremidade (+) é igual a que sai na (-)
 é igual a que sai na (-)")
27
Estabilidade verificada no Ponto de Checagem da transição da Metáfase - Anáfase

28
Ponto de checagem do fuso
Os cinetocoros que ainda nau receberam a ligacao com os microtubulos recebem um complexo proteico contendo, dentre outras proteinas as proteínas Mad2 e BubR1, que recrutam a proteina CDC20 ( ativadora do complexo promotor da anafase – APC) para esse local, formando o chamado complexo de checagem. Estrando aprisionada no complexo de checagem, CDC20 não é capaz de ativar o APC, bloqueando, desse modo,o inicio da anáfase. Quando ocorre a ligacao de microtubulos ao cinetocoro, o complexo de checagem se desfaz, liberando a CDC20. Essa proteina se liga ao APC ativando-o e possibilitando a degradacao da securina, que libera a separase, permitindo o desencadeamento da anáfase. Niveis baixos de ciclina b leva a saida de mitose. [ cromossomos de condensam, EN reorganiza e a celula se divide.
 para esse local, formando o chamado complexo de checagem. Estrando aprisionada no complexo de checagem, CDC20 não é capaz de ativar o APC, bloqueando, desse modo,o inicio da anáfase. Quando ocorre a ligacao de microtubulos ao cinetocoro, o complexo de checagem se desfaz, liberando a CDC20. Essa proteina se liga ao APC ativando-o e possibilitando a degradacao da securina, que libera a separase, permitindo o desencadeamento da anáfase. Niveis baixos de ciclina b leva a saida de mitose. [ cromossomos de condensam, EN reorganiza e a celula se divide.")
29
Na súbita transição da metafase-anáfase inicia a desfosforilação de muitas proteínas que estavam fosforiladas na prófase. Após o desligamento da M-Cdk a fosfatase passa agir sem oposição.

30
Movimento dos microtúbulos
A: Despolarização dos MTs cinetocóricos; B: Mts interpolares (1) empurram os pólos que se afastam, e Mts astrais (2) puxam os pólos distanciando um em relação ao outro. Entra tubulinas na extremidade + mais do que sai.
 empurram os pólos que se afastam, e Mts astrais (2) puxam os pólos distanciando um em relação ao outro. Entra tubulinas na extremidade + mais do que sai.")
31
Microtúbulos no cinetócoro
Movimento dos microtúbulos Microtúbulos no cinetócoro

32
Proteínas motoras

33
Movimento dos microtúbulos

34
O movimento dos microtúbulos sobrepostos na anáfase
Essa micrografia mostra a redução no grau de sobreposição dos microtubulos do fuso central. Durante a mitose de um diátomo. Etapa final da anáfase.

36
Um agrupamento dinâmico composto de filamentos de actina, de filamentos de miosina II e muitas proteínas estruturais e regulatórias.

37
Citocinese animal RhoA: GTPase controla o comportamento do anel contrátil em células animais. Estimulando a polimerização da actina. Na anáfase: miosina II começa a acumular associando-se ao anel contrátil F. A Microcrafia de fluorescencia de uma ameba deslizante fixada para actina e para miosina II. Enquanto que toda miosina II visivel se redistribuiu para o anel contrátil, comente algumas das actinas o fizeram, o restante permanece no córtex da célula. o anel de contração é suficiente para dobrar um fina agulha colocada neste local. Proteinas motoras e proteinas acessórias agora ativas ajudam a formacao do anel Em células de leveduras, o local de formação do anel é escolhido antes da mitose de acordo com uma marca colocada no córtex durante um ciclo celular prévio. Um anel de proteinas chamadas septinas forma-se antes da mitose, as septinas formam um arcabouço no qual outros componentes do anel contrátil incluindo a miosina II, se agrupam. - A expansão dos microtúbulos pode ativar alguns membros da família Rho de pequenas GTPases,que estimulam a polimerização da actina. Um membro dessa familia RhoA é necessário para citocinse Cél. Interfase – os filamentos de actina e miosina são agrupados formando uma rede cortical e, em algumas células, formam grandes feixes citoplasmáticos denominados de fibras de estresse. Conforme a célula entre em mitose: esses arranjos são desfeitos, muito das filamentos de actina são reorganizados e os de miosina II são liberados. Á medida que cromátides se separam na anáfase, a miosina II começa a se acumular rapidamente, associando-se ao anel contrátil. Nas leveduras, septinas forma um anel no qual outros componentes do anel contrátil incluindo a miosina II se agrupam em G1 tardio, marcando o local da futura citocinese. Miosina II

38
Ativação do anel contrátil

39
Citocinese centrípeta
Anel contrátil Semelhante a contração do músculo liso: Ca² calmodulina ativa a cinase da cadeia leve da miosina para fosforilar a miosina II. * inserção de uma nova membrana por fusao de vesiculas intracelulares, pra compensar o aumento na superfície da área que acompanha a divisão citoplasmática. ---- *Avanço da invaginação de um sulco de clivagem de uma célula animal em divisao *o anel de contração é suficiente para dobrar um fina agulha colocada neste local. Citocinese de um ovo de sapo - microscopia de varredura Citocinese centrípeta

40
Corpo central A clivagem está quase completa, mas as células-filha ainda não estão ligadas por uma fina corda de citoplasma contendo o restante do fuso central. Resíduos do corpo central muitas vezes permanece dentro da MP de cada célula, onde pode servir como marca no córtex, que ajuda a orientar os fusos na próxima divisão celular. - Centríolo -materno de uma ou de ambas as células-filhas separa-se do seu centriolo-filho e migra para dentro do corpo central, onde permanece alguns minutos antes de retornar para uma célula filha. – Não se sabe o que o centríolo pode fazer no corpo central para desencadear o passo final da citocinese.

41
Citocinese centrífuga
Citocinese Vegetal Membrana da vesicula = MP Componentes da vesicula = PC. Restos de microtúbulos interpolares e vesículas derivadas do CG Banda pré-profásica: microtúbulos e actina Citocinese centrífuga

42
- Célula de raiz de cebola em citocinese corada com hematoxilina ferrica. Seta indica a parede celular em formação. Não há microtúbulo astral, pois não existem centrossomos em células de plantas superiores.

43
Mitose vegetal

44
Fuso sem centrossomo: organização do fuso bipolar
Bolhas revestidas de DNA Microtubulos são formados nas regioes ao redos das bolas - os motores direcionados na extremidade mais empacotam e ordenam os microtubulos - os motores direcionados na extremidade mais empurram as extremidades menos dos microtubulos para fora das bolhas e movem os microtubulos antiparalelos - motores direcionados na extremidade menos, nas areas dos polos do fuso Embriões de inseto: Sciara

45
E as organelas ? EN RE CG Peroxissomos Mitocôndria Citoesqueleto
Na prófase ocorre gradativamente a fragmentação do nucléolo, cujos componentes em parte se dispersam pelo citoplasma na forma de córpusculos de ribonucleoproteinas, e em parte associados à periferia dos cromossomos. EM/ RE/ CG : fragmentam-se em pequenas vesículas. As vesículas nuclear contêm as lâminas B, que permanecem associadas à sua membrana interna. As laminas A ficam livres no citosol. Lisossomos, mitocôndria, peroxissomos, cloroplastos = dispersas em números equivalentes pelas células. São formadas a partir da divisão e crescimento de organelas preexistentes. Mitocôndrias e cloroplastos estão normalmente presentes em número suficiente para serem herdadas com segurança; em média, seus números aproximadamente dobram uma vez a cada ciclo. Microtúbulos (citoesqueleto) tem como função orientar as organelas as células filhas. Através de proteínas motoras: Cinesinas orientam organelas próximas ao núcleo. E Dineinas orientam organelas mais próximas ao córtex celular. RE e CG se reorganizam Citoesqueleto sofre reorganização Cloroplasto CG Lisossomos
 tem como função orientar as organelas as células filhas. Através de proteínas motoras: Cinesinas orientam organelas próximas ao núcleo. E Dineinas orientam organelas mais próximas ao córtex celular. RE e CG se reorganizam. Citoesqueleto sofre reorganização. Cloroplasto. CG. Lisossomos.")
46
Mitose sem citocinese As primeiras 13 divisões nucleares ocorrem sincronizadamente sem citocinese. Ocorre formação de 6 mil núcleos, arranjados em monacamada. A – a MP estende para o interior e estrangula para rodear cada núcleo para formar células individuais num processo chamado por celularização. Os aneis contrateis se formam na superficie da célula e a MP se invagina para envolver o núcleo. B- micrococopia de fluorescência de multiplos fusos na metáfase no embrião de Drosophila antes da celularização. A maioria dos núcleos vão migrar para o córtex e a MP estende para o interior para rodear cada núcleo celularização Osteoclastos, células do músculo do coração, alguns hepatócitos.

47
Mitose Fissão binária em células procarióticas
oriC = origem de replicação única. Depende de filamentos de proteínas FtsZ : semelhante estruturalmente a tubulina. Os filamentos de FtsZ são essenciais para o recrutamento de todas as proteínas da divisão celular no local da divisão. As proteínas direcionam o crescimento para dentro da PC e da membrana levando a formação do septo que divide a célula em duas. Cromossomos determinam de alguma forma a localização do plano de divisão celular. FtsZ é semelhante a dinamina que ocupa a função da FtsZ na divisao da mitocôndria. Se agrupa dentro do anel na região equatorial. - A] micrografia de fluorescência mostrando a localização da proteína FtsZ durante a divisão binária em E.coli. A proteína dentro de um anel no centro da célula, onde ajuda a orquestrar a divisao celular. A bacteria foi modificada por engenharia genética para produzir uma forma fluorescente de proteína FtsZ fusionada com uma proteína fluorescente verde. B] cloroplasto se dividindo [ vermelho] de uma alga vermelha também fazendo uso de um anel de proteína FtsZ [ verde] para clivagem.

48
Mitose assimétrica Os movimentos do fuso são direcionados por mudanças locais no córtex da célula; as proteínas motoras ali localizadas puxam um dos pólos do fuso, via seu microtúbulo astral, para um região apropriada. células tronco têm a propriedade fundamental de divisão assimétrica, ou seja, ao mesmo tempo em que dão origem a células tecido específicas e funcionais, produzem células indiferenciada que repõe a população de células tronco. - Segregar alguns componentes para um lado da celula [ determinantes de destino ] Plano de divisão : o fuso tem que ser movido de forma controlada, para uma região apropriada. Ovogênese = corpusculos polares primário e secundário. Uma divisao celular assimetrica segregando os componentes citoplasmaticos para somente uma célula-filha. Nucleo e os corpos polares. Nematóide. C.elegans Células de plantas: morfologia dos tecidos. Ovogênese

49
Mitose Fechada [?] Mitose aberta Mitose fechada Mitose aberta
Dinoflagelados tipicos: (protistas flagelados. A maior parte das espécies pertencem ao plâncton marinho (mais especificamente do fitoplâncton), mas são também comuns em água doce.) vários feixes de microtúbulos passam através de túneis no EM intacto para estebelecer a polaridade da divisão. Os cromossomos movem-se em associação com a membrana nuclear interna sem serem fixados aos feixes de microtúbulos; Quando esses fusos de microtúbulos pressionam a parte externa do EM, esse fica marcado por canais paralelos. Os cromossomos se fixam na membrana interna do EN oposto aos canais, e a segregação dos cromossomos ocorre dentro deste membrana nuclear canalizada. Assim o “fuso” extranuclear é usado para ordenar a membrana nuclear e dessa forma definir o plano de divisão. A mitose depende de um mecanismo de fixação da membrana. Os cromossomos precisam se fixar na membrana para sua segregação. - Hipermastigotos ( protista que possui milhares de flagelados, protozoários de intestinos de insetos) e alguns dinoflagelados não-usuais. Um único fuso central entre os dois pares de centriolos é formado em um túnel através do EN intacto. Os cinetocoros irmao tornam-se separados pelo crescimento da membrana nuclear antes de se ligare ao fuso. Os cromossomos são fixados pelo seus cinetócoros à membrana nuclear e interagem com os pólos do fuso via microtúbulos dos cinetocoros.( que se forma fora do EN.) que se fixam aos cromossomos dentro do EN. Leveduras e diátomos (algas castanhas) O EN permanece intacto. Os microtubulos do fuso formam-se dentro do núcleo, eles são formados pelos corpos dos pólos do fuso e associados com o EN; Um único microtubulo do cinetocoro fixa-se a cada cromossomo no pólo. Processo similar do que ocorre em células animais exceto que ocorre dentro do núcleo. Até agora, não existe explicação convicente para as plantas superiores e os animais apresentarem um complexo mitótico que necessita do controle e da dissolução reversível do EN. Mitose aberta
![Mitose Fechada [ ] Mitose aberta Mitose fechada Mitose aberta](http://slideplayer.com.br/slide/1566299/4/images/49/Mitose+Fechada+%5B+%5D+Mitose+aberta+Mitose+fechada+Mitose+aberta.jpg "Dinoflagelados tipicos: (protistas flagelados. A maior parte das espécies pertencem ao plâncton marinho (mais especificamente do fitoplâncton), mas são também comuns em água doce.) vários feixes de microtúbulos passam através de túneis no EM intacto para estebelecer a polaridade da divisão. Os cromossomos movem-se em associação com a membrana nuclear interna sem serem fixados aos feixes de microtúbulos; Quando esses fusos de microtúbulos pressionam a parte externa do EM, esse fica marcado por canais paralelos. Os cromossomos se fixam na membrana interna do EN oposto aos canais, e a segregação dos cromossomos ocorre dentro deste membrana nuclear canalizada. Assim o fuso extranuclear é usado para ordenar a membrana nuclear e dessa forma definir o plano de divisão. A mitose depende de um mecanismo de fixação da membrana. Os cromossomos precisam se fixar na membrana para sua segregação. - Hipermastigotos ( protista que possui milhares de flagelados, protozoários de intestinos de insetos) e alguns dinoflagelados não-usuais. Um único fuso central entre os dois pares de centriolos é formado em um túnel através do EN intacto. Os cinetocoros irmao tornam-se separados pelo crescimento da membrana nuclear antes de se ligare ao fuso. Os cromossomos são fixados pelo seus cinetócoros à membrana nuclear e interagem com os pólos do fuso via microtúbulos dos cinetocoros.( que se forma fora do EN.) que se fixam aos cromossomos dentro do EN. Leveduras e diátomos (algas castanhas) O EN permanece intacto. Os microtubulos do fuso formam-se dentro do núcleo, eles são formados pelos corpos dos pólos do fuso e associados com o EN; Um único microtubulo do cinetocoro fixa-se a cada cromossomo no pólo. Processo similar do que ocorre em células animais exceto que ocorre dentro do núcleo. Até agora, não existe explicação convicente para as plantas superiores e os animais apresentarem um complexo mitótico que necessita do controle e da dissolução reversível do EN. Mitose aberta.")
50
Obrigado !

Apresentações semelhantes













 até o momento em que esta mesma.>")





