Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Princípios básicos de Fisiologia do Exercício
Do slide nº1 ao 26 foi dado no dia07/03/06 Do slide nº Prof. Dr. Eduardo Kokubun Depto. Ed. Física UNESP-Rio Claro

2
Tipos de sobrecarga Regular Retangular Escada Rampa Impulso Misto
Irregular

3
I II III I: Domínio Moderado II: Domínio Intenso III: Domínio Severo

4
Incremental: carga e Borg
20 40 60 80 100 120 140 160 180 200 00:00 02:00 04:00 06:00 08:00 10:00 12:00 14:00 16:00 18:00 tempo Carga (w) 2 4 6 8 10 12 14 16 18 Borg
 Borg.")
5
Incremental: FC e VE 20 40 60 80 100 120 140 160 180 200 00:00 02:00 04:00 06:00 08:00 10:00 12:00 14:00 16:00 18:00 tempo FC (bpm) 10 30 50 70 90 VE (l/min) HR VE STPD
 VE (l/min) HR. VE STPD.")
6
Incremental: VO2 e VCO2 0,5 1 1,5 2 2,5 3 3,5 4 00:00 02:00 04:00
0,5 1 1,5 2 2,5 3 3,5 4 00:00 02:00 04:00 06:00 08:00 10:00 12:00 14:00 16:00 18:00 VO2 VCO2

7
Incremental: Equivalentes Respiratórios

10
Retangular: FC

11
Retangular: VE

12
Retangular: VO2

13
Retangular: VCO2

14
Retangular: RTR

15
Contribuição aeróbia e anaeróbia
Tempo Tempo VO VO 2 2 VO2SS VO2SS DO2 DO2 % % Anaer Anaer % % Aer Aer (min) (min) ( ( l/min l/min ) ) Repouso Repouso 0,40 0,40 1 1 1,63 1,63 3,75 0,02 0,12 0,46 0,59 2,12 0,0% 0,5% 3,2% 12,3% 15,7% 56,5% 100,0% 99,5% 96,8% 87,7% 84,3% 43,5% 2 2 3,16 3,16 3 3 3,29 3,29 4 4 3,63 3,63 5 5 3,73 3,73 6 6 3,75 3,75 7 7 3,75 3,75 total total 22,94 26,25 3,31 12,6% 87,4%
 (min) ( ( l/min. l/min. ) ) Repouso. Repouso. 0,40. 0, ,63. 1,63. 3,75. 0,02. 0,12. 0,46. 0,59. 2,12. 0,0% 0,5% 3,2% 12,3% 15,7% 56,5% 100,0% 99,5% 96,8% 87,7% 84,3% 43,5% ,16. 3, ,29. 3, ,63. 3, ,73. 3, ,75. 3, ,75. 3,75. total. total. 22,94. 26,25. 3,31. 12,6% 87,4%")
16
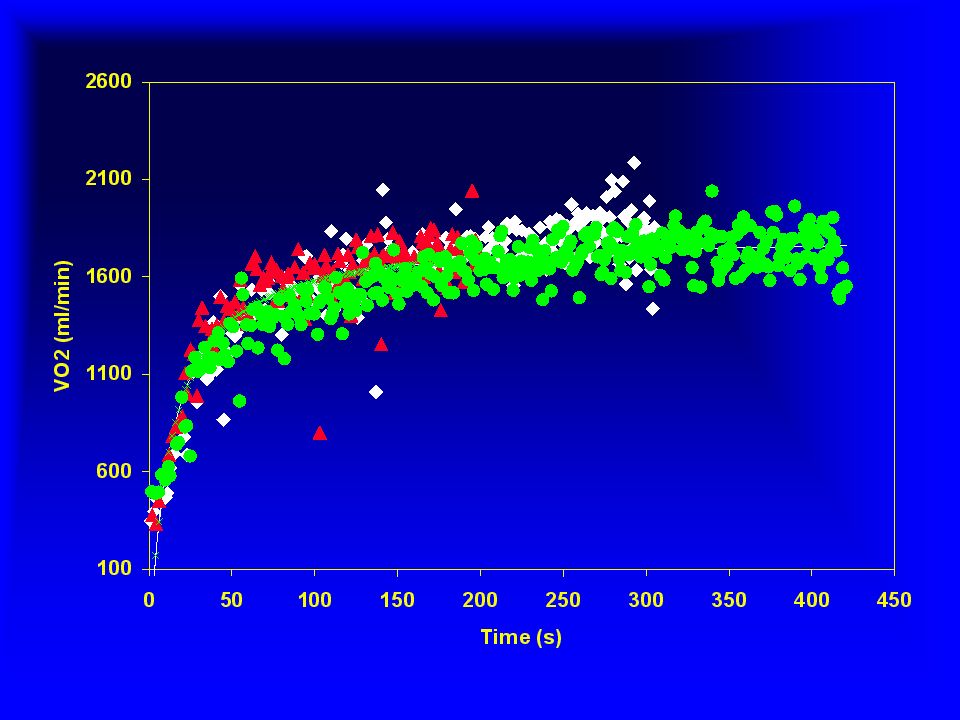
Severo (ml/min) Intenso VO Moderado Tempo (min) 2 4 6 8 10 500 1000
2 4 6 8 10 500 1000 1500 2000 2500 3000 Moderado Intenso Severo VO (ml/min) Tempo (min)
 Tempo (min)")
17
2 4 6 8 10 12 Severo Intenso Moderado Lactato (mM) Tempo (min)
 Tempo (min)")
18
O que controla a velocidade de uma reação ?
Ação de massas Catálize enzimática E As reações no organismo são regidos por dois mecanismos primários: 1) concentrações de substratos e produtos; 2) atividade catalítica da enzima. Podemos comparar o fluxo de substâncias numa reação a um sistema de caixas d'água, interligados através de um tubo com um registro Se elevarmos o nível de água na caixa S, a água tenderá a passar pelo turo em direção à caixa P. Quanto maior for o desnível entre S e P, maior será a velocidade de passagem da água. Algumas reações estão acopladas a outras. Isto significa que quando a primeira reação ocorre, parte da energia química pode ser transferido a outra substância ou a outro processo. Podemos controlar o fluxo de água também abrindo ou fechando o registro. Porém, se os níveis nas caixas S e P estiverem equilibrados, não haverá mais fluxo de água entre as caixas, independente da posição do registro. Pela analogia vemos que a relação entre os níveis na caixa d'água determina como a passagem será realizada. Do mesmo modo, em sistemas químicos a reação tende a ocorrer no sentido da substância mais concentrada para a menos concentrada. O registro equivale nesta analogia à atividade catalítica de uma enzima. Com o registro fechado, por maior que seja o desnível entre as caixas, não há fluxo de água. Do mesmo modo, a enzima inibida impede que a reação ocorra. Um sinal externo, no caso a vontade de uma pessoa em fazer a água fluir, ou um sinal biológico, tais como hormônios, presença de moduladores enzimáticos, é o estímulo ativo para o efetivo controle de uma reação. S P
 concentrações de substratos e produtos; 2) atividade catalítica da enzima. Podemos comparar o fluxo de substâncias numa reação a um sistema de caixas d água, interligados através de um tubo com um registro. Se elevarmos o nível de água na caixa S, a água tenderá a passar pelo turo em direção à caixa P. Quanto maior for o desnível entre S e P, maior será a velocidade de passagem da água. Algumas reações estão acopladas a outras. Isto significa que quando a primeira reação ocorre, parte da energia química pode ser transferido a outra substância ou a outro processo. Podemos controlar o fluxo de água também abrindo ou fechando o registro. Porém, se os níveis nas caixas S e P estiverem equilibrados, não haverá mais fluxo de água entre as caixas, independente da posição do registro. Pela analogia vemos que a relação entre os níveis na caixa d água determina como a passagem será realizada. Do mesmo modo, em sistemas químicos a reação tende a ocorrer no sentido da substância mais concentrada para a menos concentrada. O registro equivale nesta analogia à atividade catalítica de uma enzima. Com o registro fechado, por maior que seja o desnível entre as caixas, não há fluxo de água. Do mesmo modo, a enzima inibida impede que a reação ocorra. Um sinal externo, no caso a vontade de uma pessoa em fazer a água fluir, ou um sinal biológico, tais como hormônios, presença de moduladores enzimáticos, é o estímulo ativo para o efetivo controle de uma reação. S. P.")
19
ATP ADP CP Cr ATP: via final comum Glicólise e oxidação
A reconversão de ATP a ADP se faz através de três sistemas metabólicos. A primeira delas consiste na doação de um grupamento fosfato da creatina fosfato ao ADP. A segunda e a terceira vias, envolvem a hidrólise de moléculas compostas, tal como a glicose, através da glicólise incompleta ou completa. Esses sistemas incluem aquelas realizadas anaerobia ou aerobiamente.

20
Ciclo ATP A T P D CP Lactato Glicogênio Glicose Glicólise Oxidação
Ciclo de Krebs O2 CO2

21
Hidrólise do ATP: ATPase
H2O + ADP Pi + ATPase A hidrólise do ATP se inicia com o aumento da atividade da ATPase. O potencial de ação que desencadeia a contração muscular eleva a concentração de cálcio citoplasmático. Este cálcio, por sua vez, desencadeia eventos que culminam com a ativação da ATPase e a hidrólise do ATP a ADP, liberando energia para contração. Enegia

22
Reação da Creatina Fosfato Quinase (CPK)
ADP CP + ATP Cr + CPK A CP reage reversivelmente com o ADP sob a ação da enzima creatina fosfato quinase (CPK), formando ATP e creatina. Durante o exercício, a hidrólise do ATP diminui a concentração deste nucleotídeo e aumenta a do ADP. Isto ocorrendo, há tendência da reação ocorrer da esquerda para a direita, por simples ação de massas. De fato, a atividade da CPK é alta o suficiente para não limitar a velocidade desta reação. Contudo, estoques de CP no músculo são relativamente baixos, de modo que em menos de 10 s de exercício intenso, a CP pode esgotar.
, formando ATP e creatina. Durante o exercício, a hidrólise do ATP diminui a concentração deste nucleotídeo e aumenta a do ADP. Isto ocorrendo, há tendência da reação ocorrer da esquerda para a direita, por simples ação de massas. De fato, a atividade da CPK é alta o suficiente para não limitar a velocidade desta reação. Contudo, estoques de CP no músculo são relativamente baixos, de modo que em menos de 10 s de exercício intenso, a CP pode esgotar.")
23
Reação da Mioquinase (MK)
ADP + ATP AMP + MK A mioquinase (MK) é uma enzima que cataliza a reação entre duas moléculas de ADP, formando uma de ATP e outra de AMP. Esta reação além de aumentar a concentração de ATP, diminui a de ADP, contribuindo para manter o desequilíbrio dessas substâncias. O AMP formado na reação da MK pode se convertido a IMP (inosina monofosfato), com perda de uma molécula de amônia. Esta reação atua como se estivesse escoando a água da caixa P (de produtos). Em geral, admite-se que as reações da CPK e MK são vias importantes para exercícios de até 4 segundos. Após esse período, a glicólise anaeróbia começa a predominar. NH3 IMP
 é uma enzima que cataliza a reação entre duas moléculas de ADP, formando uma de ATP e outra de AMP. Esta reação além de aumentar a concentração de ATP, diminui a de ADP, contribuindo para manter o desequilíbrio dessas substâncias. O AMP formado na reação da MK pode se convertido a IMP (inosina monofosfato), com perda de uma molécula de amônia. Esta reação atua como se estivesse escoando a água da caixa P (de produtos). Em geral, admite-se que as reações da CPK e MK são vias importantes para exercícios de até 4 segundos. Após esse período, a glicólise anaeróbia começa a predominar. NH3. IMP.")
24
Ciclo ATP A T P D CP Glicogênio Glicólise Lactato Glicose Oxidação
Ciclo de Krebs O2 CO2

25
Glicólise: principais reações
Glicogênio Fosforilase HK Glicose Glicose-6-fosfato Frutose-6-fosfato PFK Frutose 1,6 difosfato Denomina-se glicólise uma série de reações que convertem a glicose-6-fosfato em piruvato. A glicose-6-fosfato pode ser produzida a partir de dois principais substratos: 1) o glicogênio armazenado no músculo, que é fosforilado a glicose-1-fosfato pela enzima fosforilase, e convertido a glicose-6-fosfato. 2) glicose extracelular, captada por um mecanismo de difusão facilitada, mediada por uma família de moléculas conhecidas como GLUT. No músculo, em particular o transportador envolvido é o GLUT4. A glicose assim captada é fosforilada pela enzima hexoquinase (HK), formando a glicose-6-fosfato. Dentre os dois substratos, o glicogênio é mais prontamente disponível para a glicólise, de modo que nó início do exercício, este é o substrato preferencial. A glicólise prossegue com a conversão de glicose6-fosfato em frutose-6-fosfato, e este em frutose-1,6-difosfato. Esta última, catalizada pela fosfofrutoquinase (PFK) é essencialmente irreversível e considerada a etapa mais importante da glicólise. A PFK sofre modulação alostérica de diferentes substratos envolvidos no metabolismo energético. Como moduladores positivos, que aceleram a reação, podem-se destacar o ADP e alcalinidade. ATP, citrato e acidez tendem a inibir a atividade da PFK. O ponto final da glicólise pode ser considerada a reação da piruvatoquinase (PK) que cataliza uma reação irreversível de formação do piruvato a partir do fosfoenolpiruvato. Do ponto de vista energético, a glicólise até a formação do piruvato, ressintetiza o ATP diretamente a partir do ADP e indiretamente formando NADH2, uma coenzima extremamente importante para o fornecimento de energia. NADH Piruvato Lactato ATP LDH PDH Acetil CoA
 o glicogênio armazenado no músculo, que é fosforilado a glicose-1-fosfato pela enzima fosforilase, e convertido a glicose-6-fosfato. 2) glicose extracelular, captada por um mecanismo de difusão facilitada, mediada por uma família de moléculas conhecidas como GLUT. No músculo, em particular o transportador envolvido é o GLUT4. A glicose assim captada é fosforilada pela enzima hexoquinase (HK), formando a glicose-6-fosfato. Dentre os dois substratos, o glicogênio é mais prontamente disponível para a glicólise, de modo que nó início do exercício, este é o substrato preferencial. A glicólise prossegue com a conversão de glicose6-fosfato em frutose-6-fosfato, e este em frutose-1,6-difosfato. Esta última, catalizada pela fosfofrutoquinase (PFK) é essencialmente irreversível e considerada a etapa mais importante da glicólise. A PFK sofre modulação alostérica de diferentes substratos envolvidos no metabolismo energético. Como moduladores positivos, que aceleram a reação, podem-se destacar o ADP e alcalinidade. ATP, citrato e acidez tendem a inibir a atividade da PFK. O ponto final da glicólise pode ser considerada a reação da piruvatoquinase (PK) que cataliza uma reação irreversível de formação do piruvato a partir do fosfoenolpiruvato. Do ponto de vista energético, a glicólise até a formação do piruvato, ressintetiza o ATP diretamente a partir do ADP e indiretamente formando NADH2, uma coenzima extremamente importante para o fornecimento de energia. NADH. Piruvato. Lactato. ATP. LDH. PDH. Acetil CoA.")
26
Ciclo ATP A T P D CP Lactato Glicogênio Glicose Glicólise Oxidação
Ciclo de Krebs O2 CO2

27
Glicólise: destino do piruvato
DH NADH Lactato NAD LDH Piruvato NAD Duas etapas da glicólise são catalizadas por desidrogenases, que reduzem NAD a NADH. Essas duas reações estão próximas ao equilíbrio, de modo que alterações nas concentrações de NAD e NADH podem afetar a velocidade da reação. Diminuição na concentração de NAD e aumento na de NADH2 diminuem a velocidade da glicólise. Dois importantes mecanismos concorrem para oxidar o NADH2 a NAD, restaurando a relação entre elas: 1) reação da lactato desidrogenase: o piruvato pode ser reduzido a lactato no citoplasma, reoxidando o NADH. 2) fosforilação oxidativa (transporte de elétrons ou cadeia respiratória): envolve uma série de reações mitocondriais de oxido-redução, que liberam energia para a ressíntese do ATP e envolvem o consumo de O2. Note que em ambos mecanismos há consumo de piruvato e reoxidação de NADH2 para NAD. Assim, ambos favorecem a continuação da glicólise, graças à redução nas concentrações de produtos e aumentando a de pelo menos um substrato. Pode-se dizer que o organismo tem “preferência” pela fosforilação oxidativa. Entretanto, a velocidade máxima dessas reações é relativamente baixa, de modo que, quando a glicólise está exacerbada, há desequilíbrio entre a formação de piruvato e a sua remoção. Neste caso, a reação da lactato desidrogenase é ativada, com consequente formação de lactato. PDH Fosforilação Oxidativa Acetil CoA
 reação da lactato desidrogenase: o piruvato pode ser reduzido a lactato no citoplasma, reoxidando o NADH. 2) fosforilação oxidativa (transporte de elétrons ou cadeia respiratória): envolve uma série de reações mitocondriais de oxido-redução, que liberam energia para a ressíntese do ATP e envolvem o consumo de O2. Note que em ambos mecanismos há consumo de piruvato e reoxidação de NADH2 para NAD. Assim, ambos favorecem a continuação da glicólise, graças à redução nas concentrações de produtos e aumentando a de pelo menos um substrato. Pode-se dizer que o organismo tem preferência pela fosforilação oxidativa. Entretanto, a velocidade máxima dessas reações é relativamente baixa, de modo que, quando a glicólise está exacerbada, há desequilíbrio entre a formação de piruvato e a sua remoção. Neste caso, a reação da lactato desidrogenase é ativada, com consequente formação de lactato. PDH. Fosforilação Oxidativa. Acetil CoA.")
28
Ciclo de Krebs Piruvato Acetil CoA CoA Oxalacetato Citrato CO2 H+
Glicogênio Glicólise Oxidação Ciclo de Krebs O2 CO2 Glicose CP Lactato Oxalacetato Citrato O ciclo de Krebs se inicia com a condensação da acetil-CoA com o oxalacetato, pela enxima citrato sintase( CS), formando citrato. Este citrato percorre uma série de reações enximáticas etermina com a formação de oxalacetato, utilizado na reação da CS. Ao longo dessas reações são formadas moléculas de CO2 e elétrons, com os prótons correspondentes, os quais oxidam as coenzimas NAD e FAD. A acetil-CoA é formada durante a metabolização de carboidratos, gorduras e proteínas. O piruvato formado na glicólise sofre ação enzimática da piruvato desidrogenase (PDH), quando perde carbono na forma de CO2 e forma NADH2. A metabolização da gordura envolve uma via denominada -oxidação, que remove dois átomos de carbono dos ácidos graxos livres, formando acetil-CoA. Já as proteínas sofrem uma hidrólise (quebra) para formação de aminoácidos. Cada aminoácido apresenta uma sequência particular de metabolização até a formação de acetil-CoA ou algum intermediário do Ciclo de Krebs. CO2 H+
, formando citrato. Este citrato percorre uma série de reações enximáticas etermina com a formação de oxalacetato, utilizado na reação da CS. Ao longo dessas reações são formadas moléculas de CO2 e elétrons, com os prótons correspondentes, os quais oxidam as coenzimas NAD e FAD. A acetil-CoA é formada durante a metabolização de carboidratos, gorduras e proteínas. O piruvato formado na glicólise sofre ação enzimática da piruvato desidrogenase (PDH), quando perde carbono na forma de CO2 e forma NADH2. A metabolização da gordura envolve uma via denominada -oxidação, que remove dois átomos de carbono dos ácidos graxos livres, formando acetil-CoA. Já as proteínas sofrem uma hidrólise (quebra) para formação de aminoácidos. Cada aminoácido apresenta uma sequência particular de metabolização até a formação de acetil-CoA ou algum intermediário do Ciclo de Krebs. CO2. H+")
29
A cadeia respiratória ATP ATP ATP FADH2 FAD Q Cit c3+ H2O NAD QH2
Os elétrons (e os prótons correspondentes) das coenzimas reduzidas são transferidas a outras coenzimas da fosforilação oxidativa. O aceptor final para esses elétons e prótons é o oxigênio, com a formação de água. Em algumas dessas passagens, são liberadas grandes quantidades de energia que são utilizadas para a ressíntese do ATP. Se a fosforilação oxidativa se iniciar no NADH2 haverá três sítios de formação de ATP, contudo se envolver a FADH2 haverá apenas dois sítios. Deste modo para cada volta no ciclo de Krebs, uma molécula de acetil-CoA poderá produzir 3 NADH2 e 1 FADH2, de modo que haverá possibilidade de formar 11 ATP. Se considerarmos que uma molécula de glicose pode produzir seis moléculas de acetil-CoA, poderemos ter a formação de 33 moléculas de ATP. Além disso, na oxidação completa, 2 moléculas de NADH2 são formadas durante a glicólise, podendo acrescentar mais 6 moléculas de ATP. Esses cálculos ilustram a enorme importância da fosforilação oxidativa. 1/2 O2 NADH+H+ Q Cit c2+
 das coenzimas reduzidas são transferidas a outras coenzimas da fosforilação oxidativa. O aceptor final para esses elétons e prótons é o oxigênio, com a formação de água. Em algumas dessas passagens, são liberadas grandes quantidades de energia que são utilizadas para a ressíntese do ATP. Se a fosforilação oxidativa se iniciar no NADH2 haverá três sítios de formação de ATP, contudo se envolver a FADH2 haverá apenas dois sítios. Deste modo para cada volta no ciclo de Krebs, uma molécula de acetil-CoA poderá produzir 3 NADH2 e 1 FADH2, de modo que haverá possibilidade de formar 11 ATP. Se considerarmos que uma molécula de glicose pode produzir seis moléculas de acetil-CoA, poderemos ter a formação de 33 moléculas de ATP. Além disso, na oxidação completa, 2 moléculas de NADH2 são formadas durante a glicólise, podendo acrescentar mais 6 moléculas de ATP. Esses cálculos ilustram a enorme importância da fosforilação oxidativa. 1/2 O2. NADH+H+ Q. Cit c2+")
30
Mecanismos de Controle do Metabolismo Energético
Via metabólica Mecanismo de Controle CP Ação de Massas Glicólise PFK - ATP/ADP Vimos 3 vias básicas para a ressíntese de ATPs durante o exercício. Obviamente, a velocidade da ressíntese da ATP deve estar finamente controlada de modo a atender a exata demanda do exercício. Todas as reações relacionadas com o metabolismo energético tem como principal regulador, as concentrações de ATP e ADP, cuja proporção deve ser mantida em valores que possibilitam o aproveitamento da energia. Oxidação KGDH - ATP/ADP

31
Metabolismo no exercício: intensidade e duração
Alta Curta A T P D Glicogênio Glicólise Oxidação Ciclo de Krebs O2 CO2 Glicose CP Lactato Intensidade Duração Baixa Longa

32
Trabalho não aproveitado
Energia no exercício Calor Energia Química Trabalho não aproveitado Trabalho Interno Trabalho Externo Útil

33
Calorimetria indireta de circuito aberto
Princípio: calorias fornecidas por combustão de oxigênio depende do nutriente Medida através do consumo de oxigênio e assim avaliar a intensidade metabólica do exercício.Além disso a relação da produção de dióxido de carbono e oxigênio é usada para indicar as contribuições de dos carboidratos e gorduras para a produção de energia.

34
Equivalente calórico de carboidratos e gorduras
C6H12O6 + 6 O2 6 CO2 + 6 H2O QR = VCO2/VO2 = 6/6 = 1 1 g CHO produz 4 Kcal 1 l O2 produz 5,05 Kcal Gorduras C16H32O O2 16 CO H2O QR =VCO2/VO2 = 16/23 = 0,70 A razão entre a produção de dióxido de carbono e o consumo de oxigênio é tradicionalmente chamado de quoeficiente respiratório (QR). O QR é usado para indicar a respiração celular e portanto o VO2 e o VCO2 resultantes do catabolismo do alimento. O QR pode ser usado para calculo do gasto energético durante o exercício -1 g Gordura produz 9 Kcal - l l O2 produz 4,7 Kcal
. O QR é usado para indicar a respiração celular e portanto o VO2 e o VCO2 resultantes do catabolismo do alimento. O QR pode ser usado para calculo do gasto energético durante o exercício. -1 g Gordura produz 9 Kcal. - l l O2 produz 4,7 Kcal.")
35
Exercício: Qual o dispendio energético ?
QR % Carboidratos % Gordura Kcal/l O2 0,70 0% 100% 4,70 0,72 7% 93% 4,72 0,74 13% 87% 4,75 0,76 20% 80% 4,77 0,78 27% 73% 4,79 0,80 33% 67% 4,82 0,82 40% 60% 4,84 0,84 47% 53% 4,86 0,86 4,89 0,88 4,91 0,90 4,93 0,92 4,96 0,94 4,98 0,96 5,00 0,98 5,03 1,00 5,05 Dados Duração 4 min Expirometria em estado estável VO2=2 l/min VCO2=1,92 l/min Calcule o QR Quantas Kcal nos 4 min ? Qual a proporção de energia de CHO ? E de gordura ? Exercitar

36
Equivalente metabólico: EM ou MET
Ocupacionais Escrever Digitar Tocar instrumento musical Assentar tijolos Carpintaria Escavar Recreacionais Dirigir automóvel Equitação, caminhada Volei recreacional Dança de salão Basquetebol Handebol, frescobol Condicionamento Caminhada 4 km/h Exercícios calistênicos Trabalho doméstico Costurar Varrer Esfregar, em pé Lavar roupas pequenas Limpar janela Cuidados pessoais Repouso Trocar de roupa Tomar banho Descer escada 1,5 2,0 2,5 3,5 5,5 7,5 3,0 4,5 9,0 10,0 1,0 Em repouso: 1 MET = 3,5 ml O2/ kg/min. Dispêndio/h = MET x Peso corporal em kg Ex: Caminhada a 4 km/h, 70 kg DE=3 x 70 = 210 Kcal/h

Apresentações semelhantes

 – Lápides 1, 2, 3» «nomes gravados, 21 de Agosto de 2008» «Ultramar.TerraWeb»>")






. Nenhuns direitos reservados, excepto para fins comerciais. Por favor, não coloque.>")











