Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Glicólise oxidação de glicose a piruvato

2
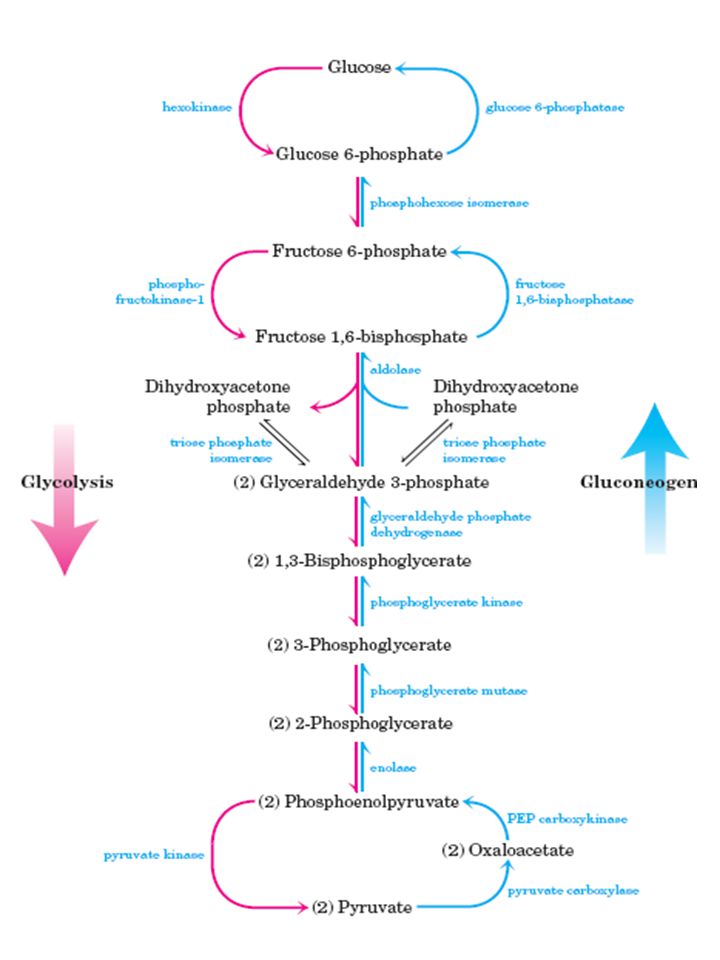
Via glicolítica

3
hexoquinase G0’ = - 16,7 kJ/mol fosfoglicose isomerase G0’ = + 1,7 kJ/mol fosfofrutoquinase-1 G0’ = - 14,2 kJ/mol aldolase G0’ = + 23,8 kJ/mol triose fosfato isomerase G0’ = + 7,5 kJ/mol gliceraldeído 3-fosfato desidrogenase G0’ = + 6,3 kJ/mol fosfoglicerato quinase G0’ = - 18,8 kJ/mol fosfoglicerato mutase G0’ = + 4,4 kJ/mol enolase G0’ = + 7,5 kJ/mol piruvato quinase G0’ = - 31,4 kJ/mol

5
1ª etapa – fase preparatória: dupla fosforilação da glicose à custa de 2 ATP

6
Reação da aldolase

7
2ª etapa: clivagem da hexose produzindo 2 trioses fosforiladas, que são interconvertíveis

8
3ª etapa – fase de pagamento: oxidação e nova fosforilação das trioses fosfato (por Pi), formando 2 moléculas de 1 intermediário com 2 grupos fosfato A oxidação do carbono torna a entrada do Pi favorável ... 1. Oxidação do aldeído (gliceraldeído 3-fosfato) a Ac. Carboxílico, com redução de NAD. Reação termodinamicamente favorável. 2 R – CO – H + 2 NAD+ + 2 H2O R – CO – OH + 2 NADH + 2 H+ 2. Ligação do Ác. Carboxílico com Pi, formando anidrido carboxílico-fosfórico, que é endergônica. 2R – CO – OH + 2 HPO R – CO – O – PO3-2 + H20 As reações ocorrem acopladas por um intermediário rico em energia. Reação pode ser inibida pelo arseniato que compete com o fosfato
 a Ac. Carboxílico, com redução de NAD. Reação termodinamicamente favorável. 2 R – CO – H + 2 NAD+ + 2 H2O 2 R – CO – OH + 2 NADH + 2 H+ 2. Ligação do Ác. Carboxílico com Pi, formando anidrido carboxílico-fosfórico, que é endergônica. 2R – CO – OH + 2 HPO4-2 2 R – CO – O – PO3-2 + H20. As reações ocorrem acopladas por um intermediário rico em energia. Reação pode ser inibida pelo arseniato que compete com o fosfato.")
9
oxidação carboximetilcisteina

10
4ª etapa: transferência dos grupos fosfato para ADP, formando 4 ATP e 2 piruvato.
Acoplamento das reações GAPDH e PGK: GAP + Pi + NAD ,3-BPG + NADH DG 0’ = +6.3 kJ/mol 1,3-BPG + ADP PG + ATP DG 0’ = kJ/mol DG 0’ = kJ/mol

11
Equação geral da glicólise:
Reaçaõ de desidratação Duas partes: - ADP ataca a fosforila do PEP formando ATP e enolpiruvato - tautomerização do PEP a piruvato Acoplamento das reações: kJ/mol suficiente para impulsionar a síntese do ATP. Equação geral da glicólise: Glicose + 2 ADP + 2 Pi + 2 NAD Piruvato + 2 ATP + 2H2O + 2NADH + 2H+

12
Como os açúcares que ingerimos na alimentação entram na via glicolítica?

13
Os destinos do piruvato

14
Destinos do piruvato em anaerobiose
Fermentação alcoólica Fermentação lática Lactato é um “bico sem saída”

15
Louis Pasteur 1861: crescimento de leveduras, por grama de glicose, maior na presença do que na ausência de ar. Glicose consumida mais lentamente na presença de ar do que na ausência. Teoria vitalista (“força vital”) Eduard Buchner 1907 – Prêmio Nobel Derruba a Teoria vitalista – a fermentação ocorre sem vida organizada – Zimases. Harden e Young 1909: isolamento do primeiro intermediário da via glicolítica. 1929: Arthur Harden - Prêmio Nobel Descoberta de um procedimento para acelerar a fermentação: adição de Pi ao meio.
 Eduard Buchner – Prêmio Nobel. Derruba a Teoria vitalista – a fermentação ocorre sem vida organizada – Zimases. Harden e Young. 1909: isolamento do primeiro intermediário da via glicolítica. 1929: Arthur Harden - Prêmio Nobel. Descoberta de um procedimento para acelerar a fermentação: adição de Pi ao meio.")
16
Via glicolítica

17
Otto Meyerhoff 1922: Prêmio Nobel
Descoberta da correlação entre o consumo de oxigênio e o metabolismo do ácido lático nos músculos de coelho. Ativador: obtido por autólise de levedura. O ativador perde a atividade se aquecido por 1 minuto a 50 ºC e conserva-se bem em gelo. Você pode imaginar a natureza desse ativador? Para você é espantoso que se obtenha um ativador de músculo de coelho a partir de levedura?

18
Metabolismo do Etanol no fígado:
ADH Álcool desidrogenase Ressaca ALDH Acetaldeído desidrogenase Hipoglicemia

19
Sensibilidade diferencial ao álcool
Consumo de álcool segundo diferentes padrões levou a uma evolução divergente. Existem várias enzimas ADH no homem: dímeros (5 genes). ADH são essenciais pois quebram e metabolizam as moléculas de álcool (tóxico) que é absorvida para o sangue. População do Sudeste Asiático: maior intolerância ao álcool – acúmulo de acetaldeído – rubor alcoólico (“Asian flush”) Alcoolismo (tolerância ao álcool) Populações europeias:alleles ADH2 e ADH3 menos ativas metabolizam lentamente o etanol Intolerância ao álcool: - Sudeste asíatico: ~ 50 % pop. possui o allele mutante ALDH2*2 (8% da atividade do gene wt)
. ADH são essenciais pois quebram e metabolizam as moléculas de álcool (tóxico) que é absorvida para o sangue. População do Sudeste Asiático: maior intolerância ao álcool – acúmulo de acetaldeído – rubor alcoólico ( Asian flush ) Alcoolismo (tolerância ao álcool) Populações europeias:alleles ADH2 e ADH3 menos ativas metabolizam lentamente o etanol. Intolerância ao álcool: - Sudeste asíatico: ~ 50 % pop. possui o allele mutante ALDH2*2 (8% da atividade do gene wt)")
20
Via glicolítica G0’ = - 31,4 kJ/mol G0’ = - 16,7 kJ/mol

21
Hexoquinase: Glicose + ATP Glicose 6-fosfato +ADP + H+ HK
Isoformas I, II e III: cinética michaeliana com Km < 0,1 mM, ou seja, funcionam sempre em Vmáx. [glicose] plasm = 5 a 8 mM HK inibidor

22
Glicoquinase e Hexoquinase
Hexoquinase (músculo): I, II, e III Glicoquinase ou Hexoquinase IV – presente no fígado: menor afinidade pela glicose. Ligada a uma proteína reguladora forma um complexo inativo. Hexoquinase IV é regulada pelo nível de glicose no sangue: regulação por seqüestro no núcleo celular Após refeição Fígado não compete com demais órgãos pela glicose escassa. Durante jejum Vindo da gliconeogênese hepatócito
: I, II, e III. Glicoquinase ou Hexoquinase IV – presente no fígado: menor afinidade pela glicose. Ligada a uma proteína reguladora forma um complexo inativo. Hexoquinase IV é regulada pelo nível de glicose no sangue: regulação por seqüestro no núcleo celular. Após refeição. Fígado não compete com demais órgãos pela glicose escassa. Durante jejum. Vindo da gliconeogênese. hepatócito.")
23
HEXOQUINASE IV Glicoquinase (Hexoquinase IV) não é inibida por glicose 6-fosfato e tem maior Km pela glicose. É importante no fígado para garantir que glicose não seja desperdiçada quando estiver abundante, sendo encaminhada para síntese de glicogênio e ácidos graxos. Além disso, quando a glicose está escassa, garante que tecidos como cérebro e músculo tenham prioridade no uso

24
PFK-1 Fru 6-F Fru 1,6-bF a partir desse ponto o açúcar está comprometido com a via glicolítica Reação altamente exergônica e irreversível DG0’ = - 14,2 kJ/mol Além do sítio ativo essa enzima possui diversos sítios onde inibidores e ativadores alostéricos se ligam.

25
Fosfofrutoquinase 1 (PFK 1):
Frutose 6-fosfato + ATP Frutose 1,6-bifosfato + ADP + H+ Efetuadores alostéricos: Negativos: ATP (retroinibição) e Citrato (CK) Positivos: AMP (musc. Esq.) e frutose 2,6-bifosfato
 e Citrato (CK) Positivos: AMP (musc. Esq.) e frutose 2,6-bifosfato.")
26
FRUTOSE 2,6-BISFOSFATO Em 1980, foi observado que frutose 2,6-bisfosfato ativava a fosfofrutoquinase aumentando sua afinidade pelo substrato frutose 6-fosfato. Além disso, diminuia o efeito inibitório do ATP Frutose 2,6-bisfosfato é um ativador alostérico que desloca o equilíbrio conformocional da enzima para sua forma ativa. É produzido pela FOSFOFRUTOQUINASE 2 (PFK 2).
.")
27
PFK2/FBPASE2

28
Enzima bifuncional: 6-fosfofruto-2-quinase/frutose 2,6-bifosfatase
Regulação alostérica PFK2 Ativa PFK1 + glicólise Regulação por controle covalente: substrato para PKA

29
Piruvato Quinase: PEP + ADP Piruvate + ATP = ativação anterógrada
Regulação alostérica = ativação anterógrada Regulação por controle covalente

30
PIRUVATO QUINASE Último passo da via glicolítica. Fluxo de saída.
Produz ATP e Piruvato. Também é um tetrâmero presentando diferentes isoformas em diferentes tecidos. Isoforma L (fígado) e isoforma M (músculo). Muitas propriedades em comum: - Frutose 1,6-bisfosfato: ativa - ATP: inibe alostericamente - Alanina: produzida a partir de piruvato, inibe a PIK.
 e isoforma M (músculo). Muitas propriedades em comum: - Frutose 1,6-bisfosfato: ativa. - ATP: inibe alostericamente. - Alanina: produzida a partir de piruvato, inibe a PIK.")
31
No entanto, as isoformas L (fígado) e M (músculo) diferem na regulação por modificação covalente: fosforilação. A isoforma L é inativada ao ser fosforilada quando o nível de glicose no sangue cai (estímulo disparado pelo glucagon)
")
32
Regulação da via glicolítica

33
Via antagônica a glicólise: Gliconeogênese
Síntese de glicose a partir de compostos que não são carboidratos: aminoácidos, lactato e glicerol.

34
Gliconeogênese Alguns tecidos dependem quase completamente de glicose para energia metabólica depleção de glicose gliconeogênese

36
Conversão de piruvato a fosfoenolpiruvato

37
* Músculo e cérebro não contêm esta enzima
Frutose 1,6-bisfosfato + H2O frutose 6-fosfato + Pi Frutose 1,6-bisfosfatase Glicose 6-fosfato + H2O glicose + Pi Glicose 6-fosfatase * Músculo e cérebro não contêm esta enzima

38
Fosfofrutoquinase 1 e Frutose 1,6-bifosfatase

39
Frutose-2,6- bifosfato

40
Regulação da via glicolítica
Hexoquinase X Glicoquinase Fosfofrutoquinase Piruvatoquinase

Apresentações semelhantes