Carregar apresentação
A apresentação está carregando. Por favor, espere

1
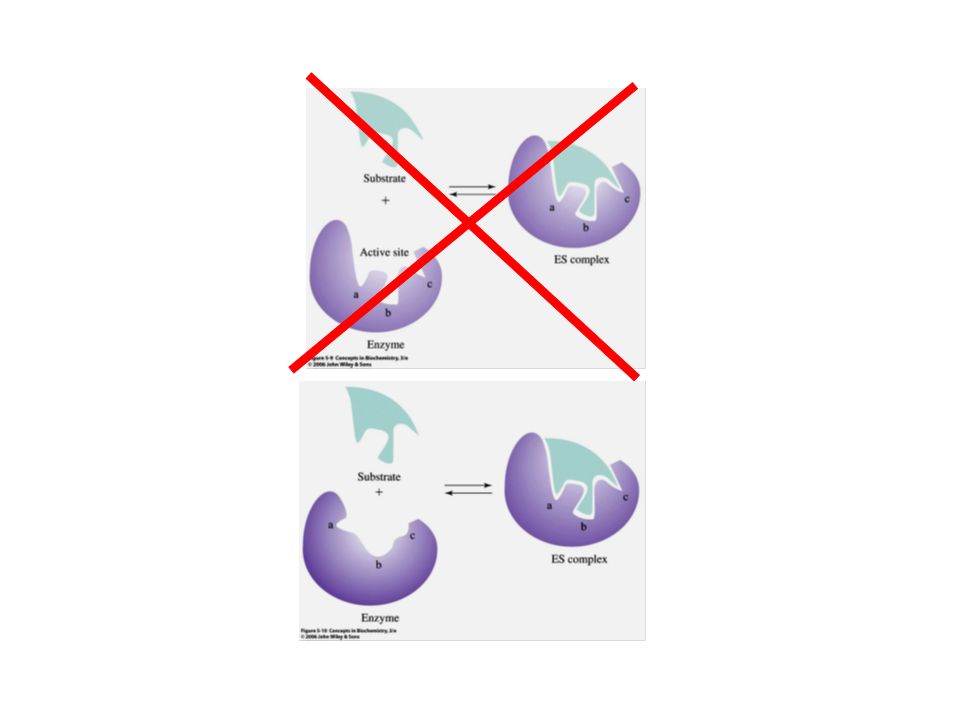
Especificidade Formação de um complexo enzima-substrato específico
Pontes de hidrogênio, interações hidrofóbicas e iônicas Depende da estrutura tridimensional Uma das características mais importantes das enzimas: especificidade. Interações hidrofóbicas, iônicas, pontes de hidrogênio estabilizam a ligação entre enzima e substrato. Essas interações afastam as moléculas de água que solvatam tanto a o sítio ativo da enzima quanto o substrato.

2
Energia de Ligação A formação de cada interação fraca no complexo ES é acompanhada da liberação de uma pequena quantidade de energia livre que estabiliza a interação Energia de ligação (ΔGB) Principal fonte de energia livre utilizada pelas enzimas para diminuição da ΔG‡
 Principal fonte de energia livre utilizada pelas enzimas para diminuição da ΔG‡")
3
Secretados como zimogênios, precursores inativos das enzimas: tripsinogênio, quimotripsinogênio, proelastase

4
Especificidade São bastante parecidas estruturalmente.
O sítio ativo contem a mesma tríade catalítica . Outros ami Resíduo de serina que dá o nome a classe de enzimas: serino proteases

5
A especificidade é dada pelo resíduo que está no bolsão S1 da enzima.
Tripsina: cliva ligações peptídicas de cadeia grande, carregados positivamente (lisina ou arginina). Resíduo no bolsão: aspartato Quimotripsina: o bolsão é mais hidrofóbico (pref. Por tirosina, fenilalanina e triptofano) Elastase: a fenda é muito menor, preferencia por alanina, glicina e valina
. Resíduo no bolsão: aspartato. Quimotripsina: o bolsão é mais hidrofóbico (pref. Por tirosina, fenilalanina e triptofano) Elastase: a fenda é muito menor, preferencia por alanina, glicina e valina.")
6
Chave e Fechadura O módulo SH2 - liga-se à tirosina fosfatada, que se adapta ao sítio ativo da enzima tal como uma chave faz a sua fechadura. Domínio SH2: domínio proteico conservado presente na Src. Permite que proteínas contendo contendo esse domínio se acoplem em outras proteínas contendo tirosinas fosforiladas.

7
Chave e Fechadura Emil Fischer, 1894
Hipótese enganadora quando aplicada a catálise enzimática Enzimas complementares seriam inibidores Complementar ao estado de transição

8
Encaixe Induzido Prevê um sítio de ligação não totalmente pré-formado, mas sim moldável à molécula do substrato; A enzima se ajustaria à molécula do substrato na sua presença

9
Encaixe Induzido

11
Grupos Catalíticos Catálise geral ácido-base
Ocorre com a participação de aminoácidos com cadeias laterais ionizáveis, capazes de doar ou liberar prótons durante a catálise para estabilizar os intermediários (H+, OH-)
")
12
Grupos Catalíticos Catálise Covalente
Resulta do ataque nucleofílico ou eletrofílico de um radical do sítio catalítico sobre o substrato, ligando-o covalentemente à enzima e induzindo a sua transformação em produto.

13
Frutose 2,6-bifosfatase

14
Grupos Catalíticos Catálise por Íon Metálico
Metais firmemente ligados ao sítio catalítico interagem entre a enzima e o substrato estabilizando o estado de transição Um grupo prostético de zinco na enzima é coordenado em três posições por cadeias laterais de histidina. A quarta posição de coordenação está ocupada pela água. Isso causa a polarização da ligação entre oxigênio-hidrogênio, fazendo o oxigênio ligeiramente mais negativa, enfraquecendo assim a ligação. Uma quarta histidina é colocado próximo ao substrato da água e recebe um próton, em um exemplo de ácido geral. Isto deixa um hidróxido ligado ao zinco. O sítio ativo também contém um bolso especificidade para o dióxido de carbono, aproximando-o do grupo hidróxido. Isto permite que o hidróxido rico em electrons ataque o dióxido de carbono, formando bicarbonato.

15
Mecanismo de Ação da Quimiotripsina
Grupos catalíticos específicos contribuindo para catálise: ácido–base, covalente Quimiotripsina possui mecanismos de catálise variados para promover a quebra do peptídeo

16
Covalente Substrato encaixa no sítio ativo, bolsão hidrofóbico -> Hist. rouba proton da Ser, tornando o oxigenio fortemente nucleofilico Ataque nucleófílico no grupo carbonil (C=O), formando um intermediário tetraédrico. Oxigênio do carbonil se torna instável e é estabilizado por ligações de hidrogênio no bolsão do oxiânion. A instabilidade do intermediário colapsa o intermediário tetraédrico, formando uma nova dupla ligação com o carbono, deslocando a ligação com o grupo amida. A água é desprotonada por catálise geral básica, gerando o OH- altamente nucleofílico Formação de um novo intermediário tetraédrico com O instavel Colapso do intermediário com formação de nova dupla ligação e liberação da ligação a serina Ac.-Base
, formando um intermediário tetraédrico. Oxigênio do carbonil se torna instável e é estabilizado por ligações de hidrogênio no bolsão do oxiânion. A instabilidade do intermediário colapsa o intermediário tetraédrico, formando uma nova dupla ligação com o carbono, deslocando a ligação com o grupo amida. A água é desprotonada por catálise geral básica, gerando o OH- altamente nucleofílico. Formação de um novo intermediário tetraédrico com O instavel. Colapso do intermediário com formação de nova dupla ligação e liberação da ligação a serina. Ac.-Base.")
17
Efeito do pH na Atividade Enzimática
Mudanças extremas de pH podem alterar a estrutura enzimática por repulsão de cargas. Mudanças mais brandas podem afetar a atividade enzimática por afetar resíduos no sítio catalítico. Glicose-6-fosfato: dois grupos ionizáveis = pH otimo proximo de 7,5 Prepsina: um grupo ionizável, atividade cai conforme pH aumenta

18
Efeito do pH na Atividade Enzimática
A ionização de aminoácidos (aa) pode provocar modificações na conformação da enzima. pH > pI pH do meio esta alcalino; concentração reduzida de íons H+ no meio; os aa liberam H+, ficando eletricamente negativo. pH < pI pH do meio esta ácido; excesso de íons H+ no meio; os aa recebem H+, ficando eletricamente positivo.
 pode provocar modificações na conformação da enzima. pH > pI. pH do meio esta alcalino; concentração reduzida de íons H+ no meio; os aa liberam H+, ficando eletricamente negativo. pH < pI. pH do meio esta ácido; excesso de íons H+ no meio; os aa recebem H+, ficando eletricamente positivo.")
19
Efeito da Temperatura na Atividade Enzimática
quimiotripsina de porco enzima isolada de um camarão do Alaska enzima isolada de uma bactéria de fontes termais Aumento da temperatura aumenta a chance de moléculas colidirem (dá energia ao sistema) Altas temperaturas – desnaturação (perda de interações não covalentes) – proteína perde a estrutura 3d
 Altas temperaturas – desnaturação (perda de interações não covalentes) – proteína perde a estrutura 3d.")
Apresentações semelhantes