Carregar apresentação
A apresentação está carregando. Por favor, espere

4
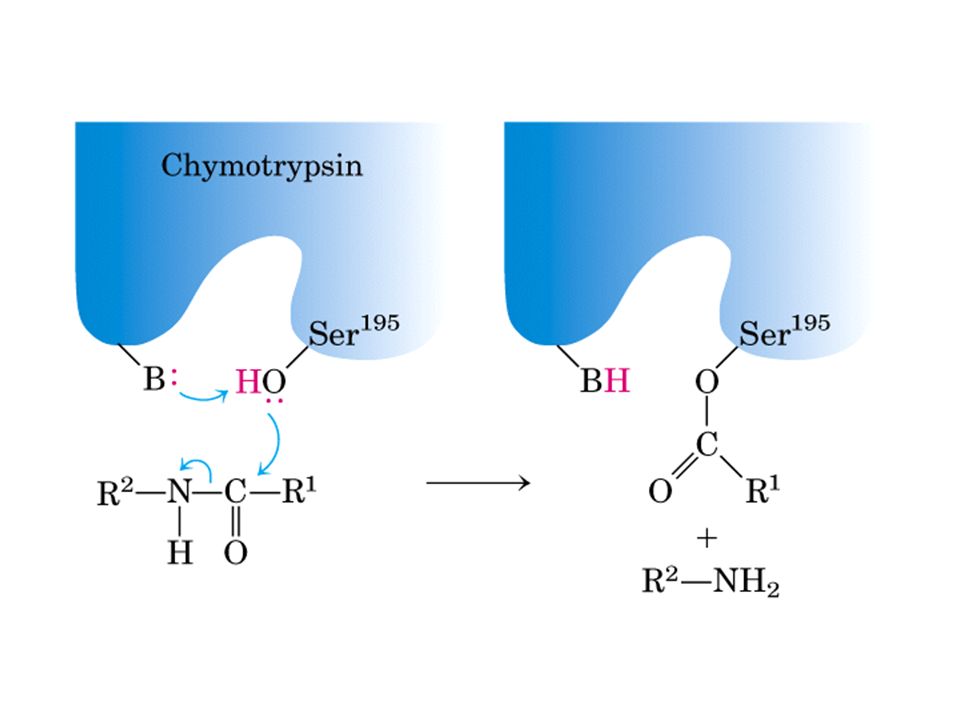
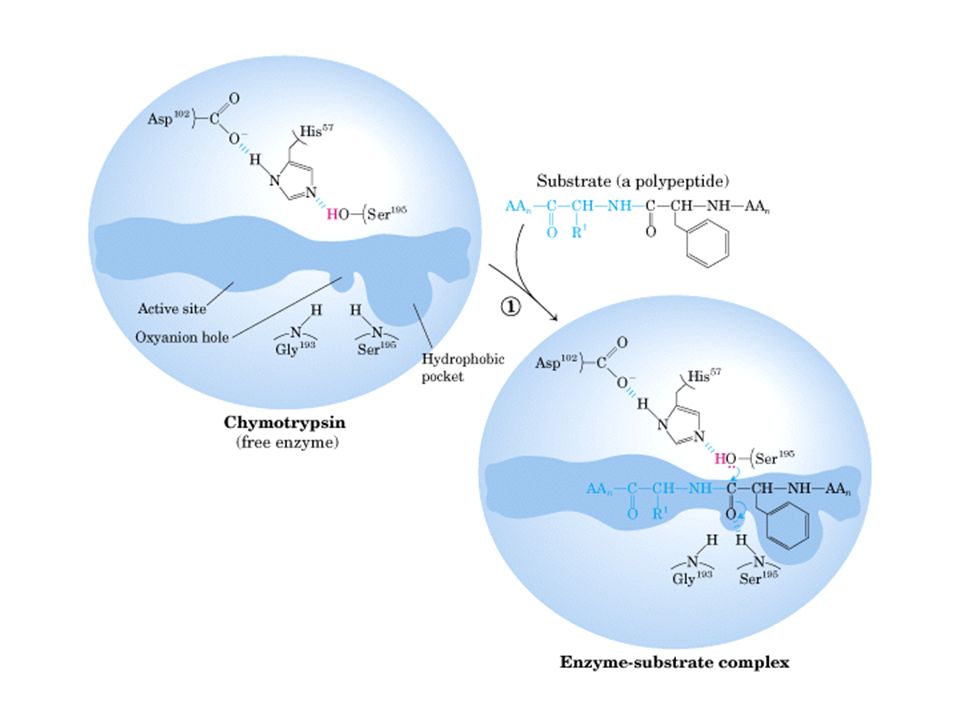
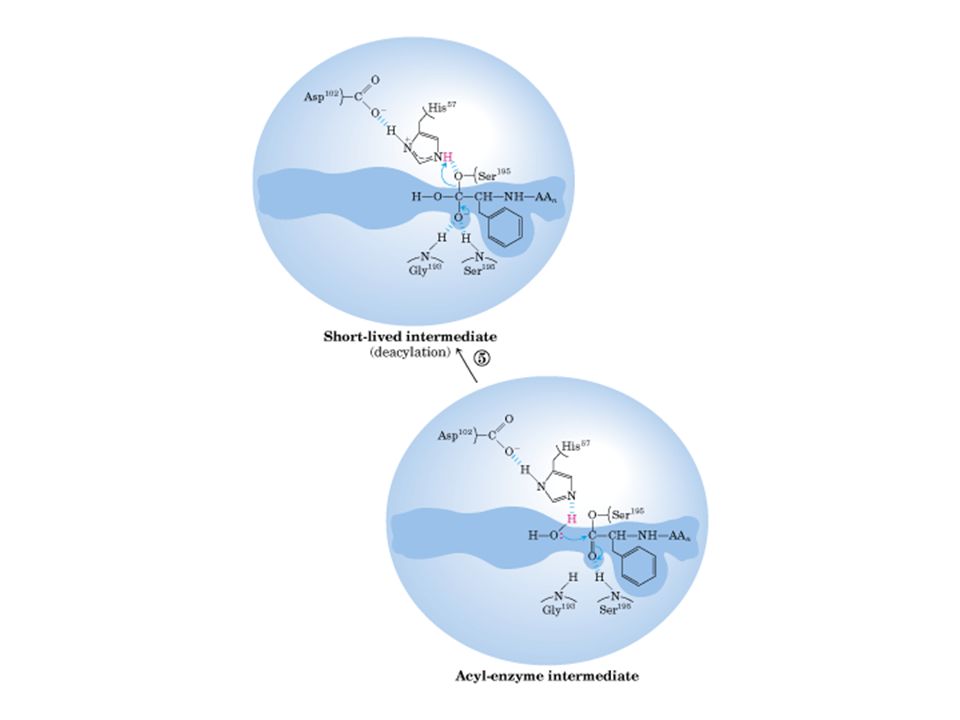
Quimotripsina

9
COMPLEMENTARIDADE ENTRE
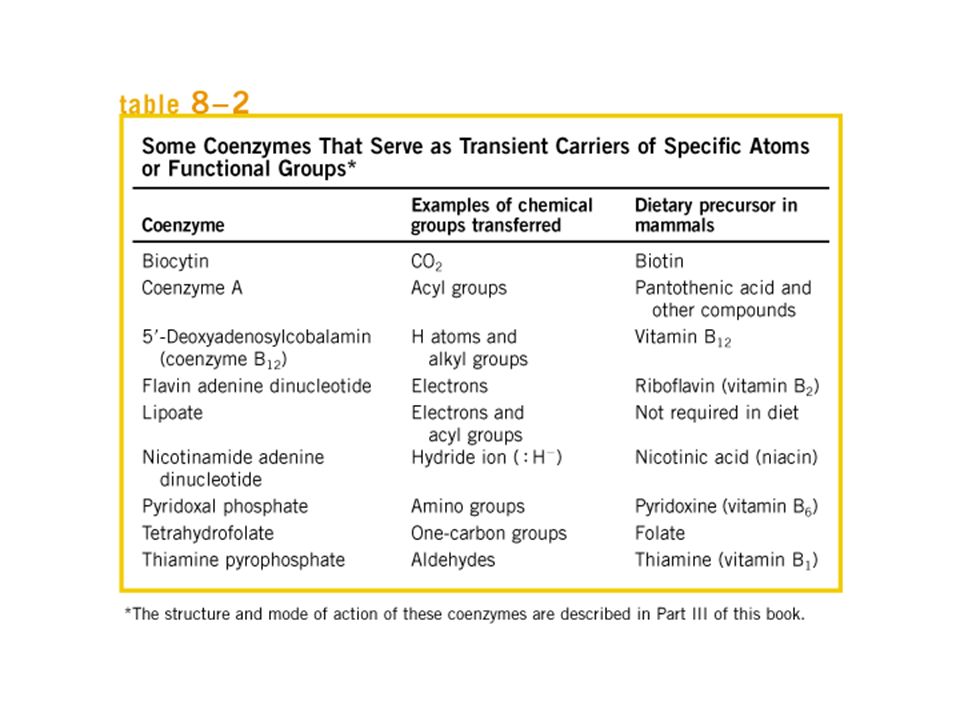
DIIDROFOLATO REDUTASE ES SEUS SUBSTRATOS Amarelo: tetraidrofolato Vermelho: NADP+

10
“BASTÃO-ASE” DGM contribuido pelas interações entra e enzima e o estado de transição

16
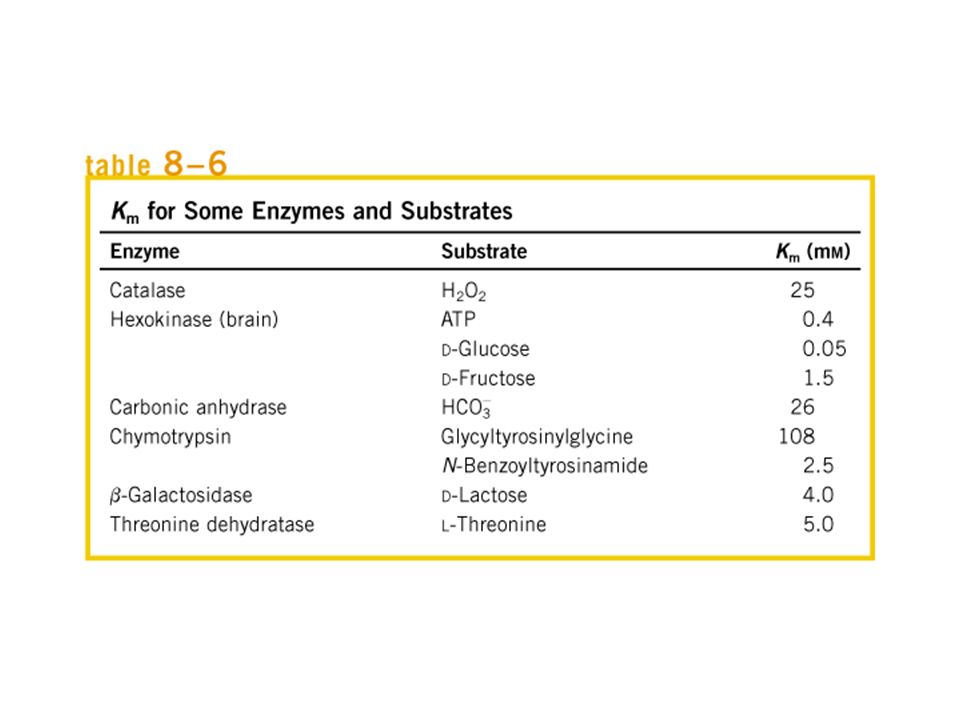
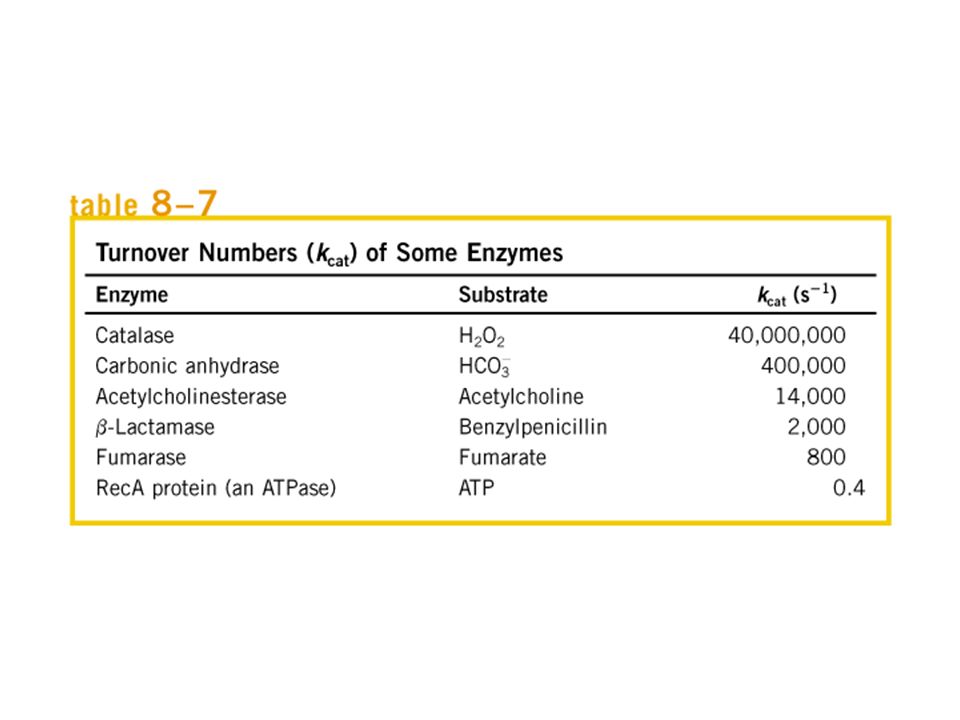
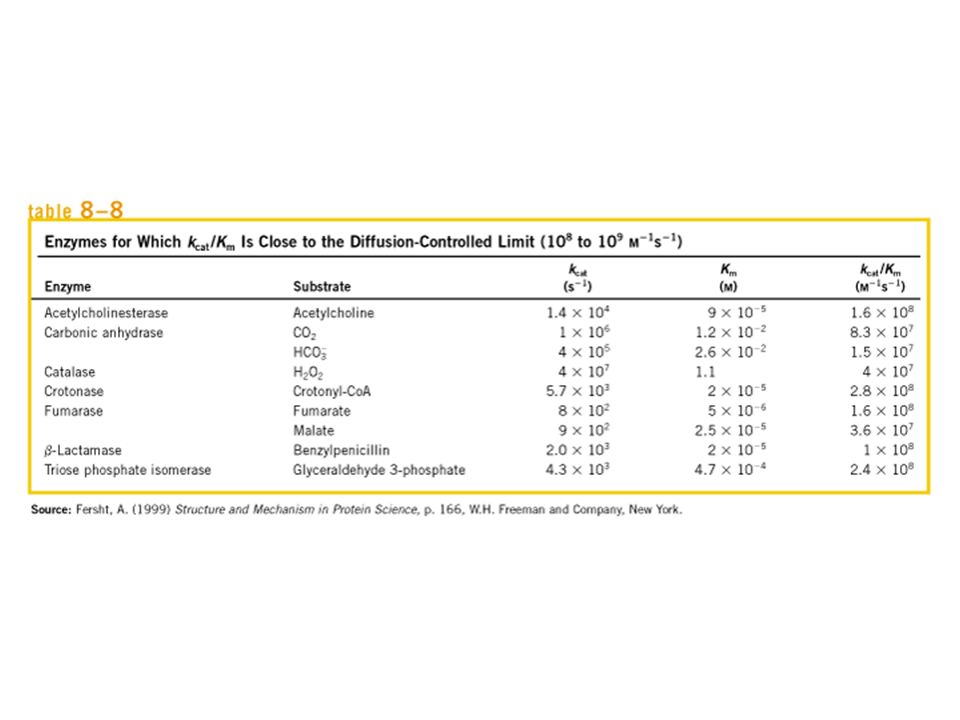
Cinêtica Enzimática Michaelis-Menten

17
k1 kcat E + S ES E + P k-1 V = kcat[ES] Assumir que: d[ES]/dt = 0
k1[E][S] – k-1[ES] – kcat[ES] = 0 [E]tot = [E] + [ES] Vmax = kcat x [E]tot KM = k-1/k1
![k1 kcat E + S ES E + P k-1 V = kcat[ES] Assumir que: d[ES]/dt = 0](http://slideplayer.com.br/slide/45971/1/images/17/k1+kcat+E+%2B+S+%EF%83%9F%EF%83%A0+ES+%EF%83%A0+E+%2B+P+k-1+V+%3D+kcat%5BES%5D+Assumir+que%3A+d%5BES%5D%2Fdt+%3D+0.jpg "k1[E][S] – k-1[ES] – kcat[ES] = 0. [E]tot = [E] + [ES] Vmax = kcat x [E]tot KM = k-1/k1.")
18
Vo = Vmax[S] = kcatx[E]totx[S]
Km+[S] Km+[S]
![Vo = Vmax[S] = kcatx[E]totx[S]](http://slideplayer.com.br/slide/45971/1/images/18/Vo+%3D+Vmax%5BS%5D+%3D+kcatx%5BE%5Dtotx%5BS%5D.jpg "Km+[S] Km+[S]")
20
Diagrama Lineweaver-Burk (Duplo-reciprocal)
")
24
INIBIÇÃO COMPETITIVO:
substrato e inibidor competem para o mesmo sítio Km kcat Vo = Vmax[S]/(aKm + [S]) a = 1 + [I]/KI
 a = 1 + [I]/KI.")
25
a’ = 1 + [I]/K’I Vo = Vmax[S]/(Km + a’[S]) Quando [S]>>Km, Vo = Vmax/a’ INIBIÇÃO NÃO-COMPETITIVO: substrato e inibidor competem para sítios diferentes; inibidor somente liga ao complexo ES
![a’ = 1 + [I]/K’I Vo = Vmax[S]/(Km + a’[S]) Quando [S]>>Km, Vo = Vmax/a’ INIBIÇÃO NÃO-COMPETITIVO:](http://slideplayer.com.br/slide/45971/1/images/25/a%E2%80%99+%3D+1+%2B+%5BI%5D%2FK%E2%80%99I+Vo+%3D+Vmax%5BS%5D%2F%28Km+%2B+a%E2%80%99%5BS%5D%29+Quando+%5BS%5D%3E%3EKm%2C+Vo+%3D+Vmax%2Fa%E2%80%99+INIBI%C3%87%C3%83O+N%C3%83O-COMPETITIVO%3A.jpg "substrato e inibidor competem para sítios diferentes; inibidor somente liga ao complexo ES.")
26
Vo = Vmax[S]/(aKm + a’[S])
![Vo = Vmax[S]/(aKm + a’[S])](http://slideplayer.com.br/slide/45971/1/images/26/Vo+%3D+Vmax%5BS%5D%2F%28aKm+%2B+a%E2%80%99%5BS%5D%29.jpg "Vo = Vmax[S]/(aKm + a’[S])")
27
Diagrama Lineweaver-Burk para inibição competitiva
a = 1 + [I]/KI

28
Diagrama Lineweaver-Burk para inibição não-competitiva

29
Diagrama Lineweaver-Burk para inibição mista

30
ativa inativa EH+ E + H+ pKa = 3,5

31
inativa ativa inativa EH22+ EH+ + H+ E + 2H+ pKa = 6, pKa = 9,0

32
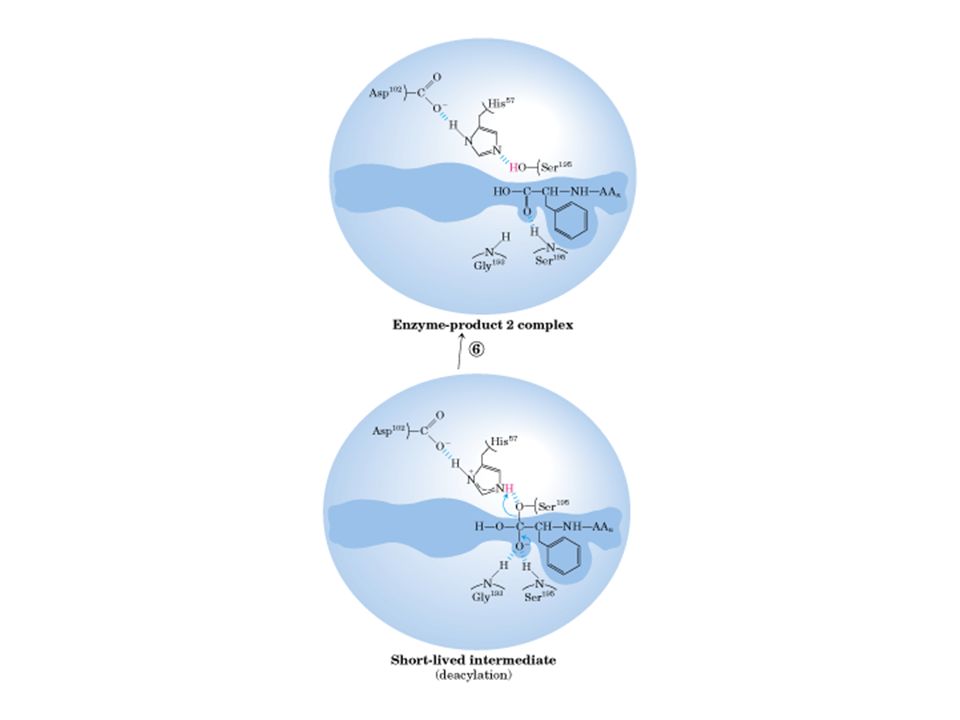
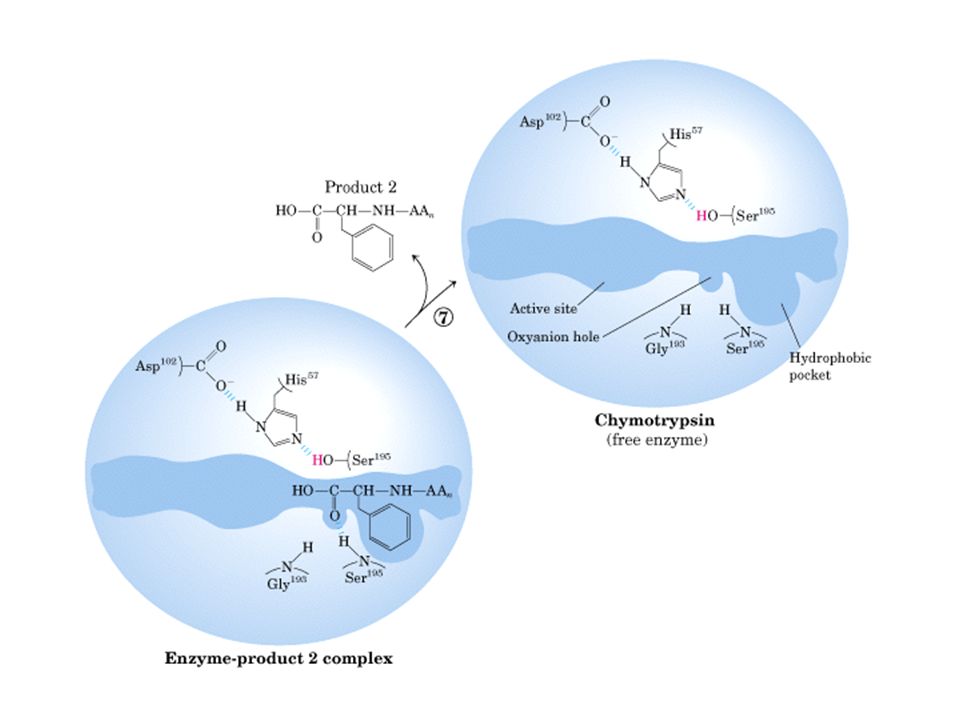
Quimotripsina

33
Quimotripsina vermelha: ser195, asp 102, his57

34
Quimotripsina vermelha: ser195, asp 102, his57

35
Sítio ativo de quimotripsina

45
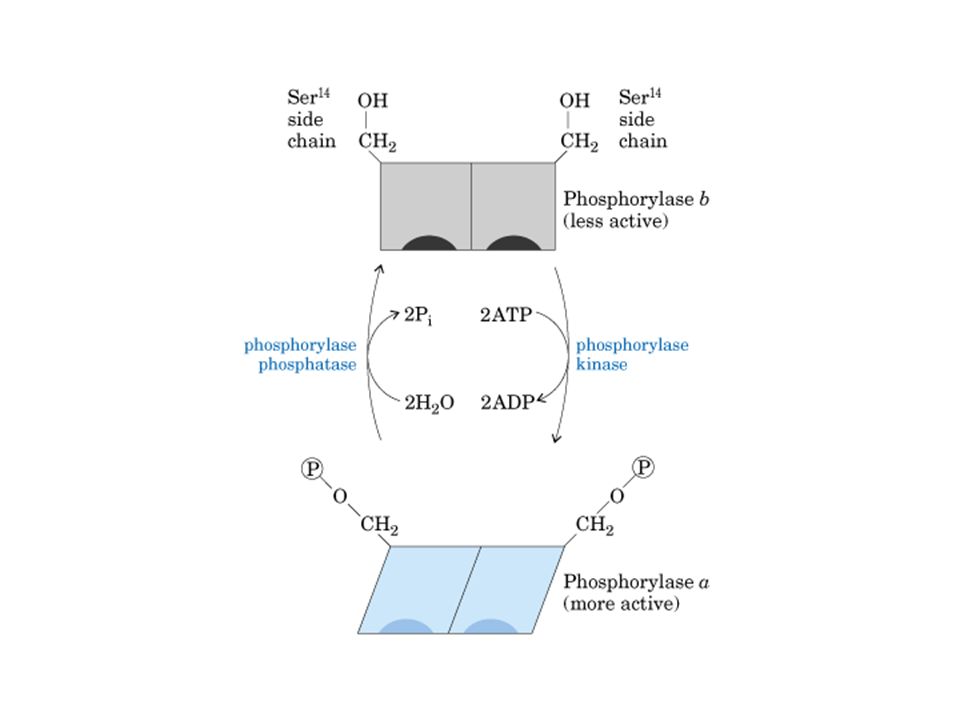
HEXOQUINASE Sem glicose Com glicose

47
Inibição “Feedback”

48
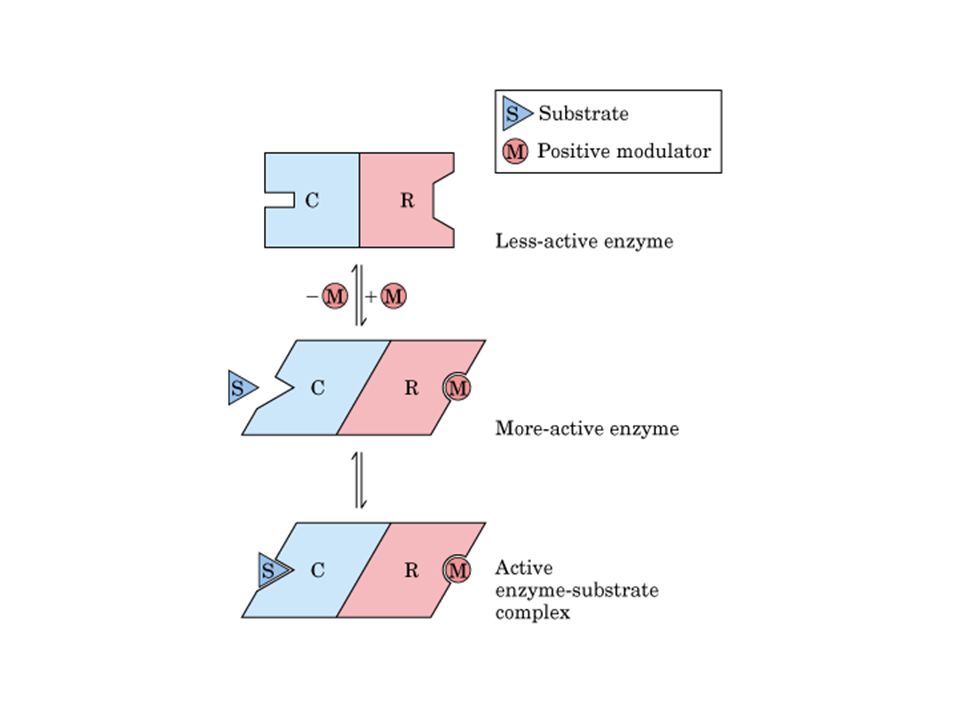
Alosteria: curva sigmoidal de uma enzima homotrópica:
o substrato age como uma moduladora positiva

49
Alosteria: curvas sigmais de uma enzima em que
moduladoras positivas e negativas modificam K0.5 sem modificar Vmax.

50
Alosteria: curvas sigmoidais de uma enzima em que
moduladoras positivas e negativas modificam Vmax Sem modificar K0.5 (menos comum).
.")
54
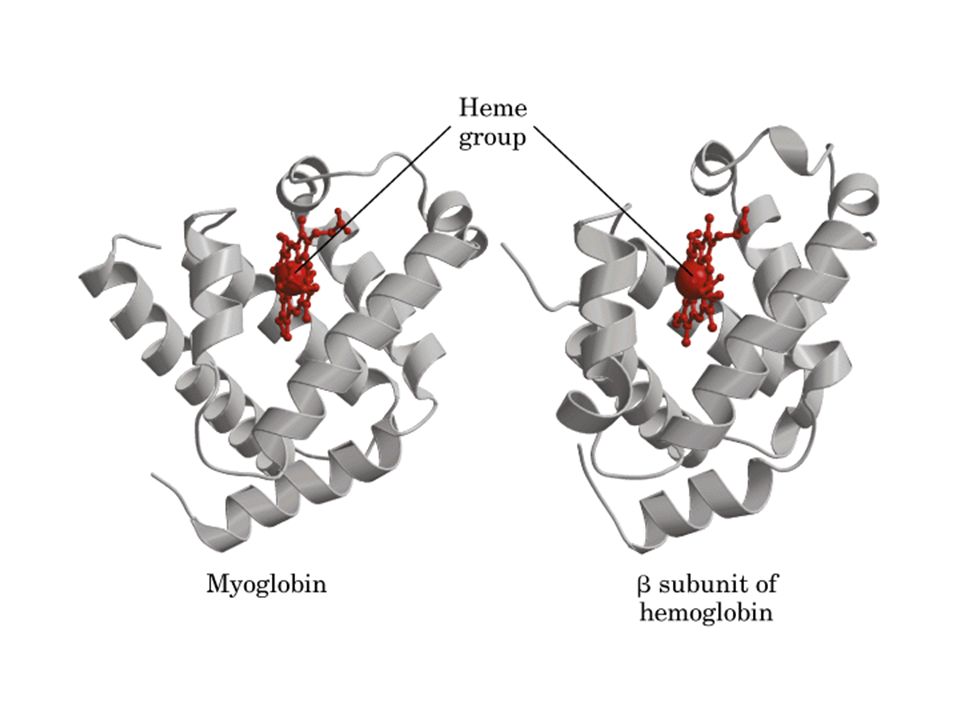
Estrutura da mioglobina (153 aminoácidos, MW = 16700 Da)
")
55
Estrutura básica das Porfirinas 4 anels de pirrole conectados por pontes de =C-

56
HEME = Prorporfirina IX

57
HEME

58
His próximal

59
P + L PL Ka = [PL]/[P][L] = 1/Kd q = [PL]/{[PL]+[P]}
![P + L PL Ka = [PL]/[P][L] = 1/Kd q = [PL]/{[PL]+[P]}](http://slideplayer.com.br/slide/45971/1/images/59/P+%2B+L+%EF%83%A0+PL+Ka+%3D+%5BPL%5D%2F%5BP%5D%5BL%5D+%3D+1%2FKd+q+%3D+%5BPL%5D%2F%7B%5BPL%5D%2B%5BP%5D%7D.jpg "P + L PL Ka = [PL]/[P][L] = 1/Kd q = [PL]/{[PL]+[P]}")
60
P + L PL Ka = [PL]/[P][L] = 1/Kd q = [PL]/{[PL]+[P]}
Ka = constante de associação, Kd = constante de dissociação = 1/Ka Fração de sítios ocupados = q q = [PL]/{[PL]+[P]} q = Ka[P][L]/{Ka[P][L]+[P]} q = Ka[L]/{Ka[L]+1} q = [L]/{[L]+1/Ka} q = [L]/{[L]+Kd]} equação tipo x = y/(y+z) descreve uma hipérbola Quando [L]=Kd=1/Ka, q = 0,5 (50% dos sítios ocupados)
![P + L PL Ka = [PL]/[P][L] = 1/Kd q = [PL]/{[PL]+[P]}](http://slideplayer.com.br/slide/45971/1/images/60/P+%2B+L+%EF%83%9F%EF%83%A0+PL+Ka+%3D+%5BPL%5D%2F%5BP%5D%5BL%5D+%3D+1%2FKd+q+%3D+%5BPL%5D%2F%7B%5BPL%5D%2B%5BP%5D%7D.jpg "Ka = constante de associação, Kd = constante de dissociação = 1/Ka. Fração de sítios ocupados = q. q = [PL]/{[PL]+[P]} q = Ka[P][L]/{Ka[P][L]+[P]} q = Ka[L]/{Ka[L]+1} q = [L]/{[L]+1/Ka} q = [L]/{[L]+Kd]} equação tipo x = y/(y+z) descreve uma hipérbola. Quando [L]=Kd=1/Ka, q = 0,5 (50% dos sítios ocupados)")
61
Curva de ligação de oxigênio a mioglobina P50 = 0,26 kPa
= fração dos sítios ocupados com ligantes = [L]/{[L]+Kd]} = [02]/{[02]+ [02] 50} =p02/{p02+P50}

62
Ligação de oxigênio a heme

63
Ligação de monôxido de carbono a heme

64
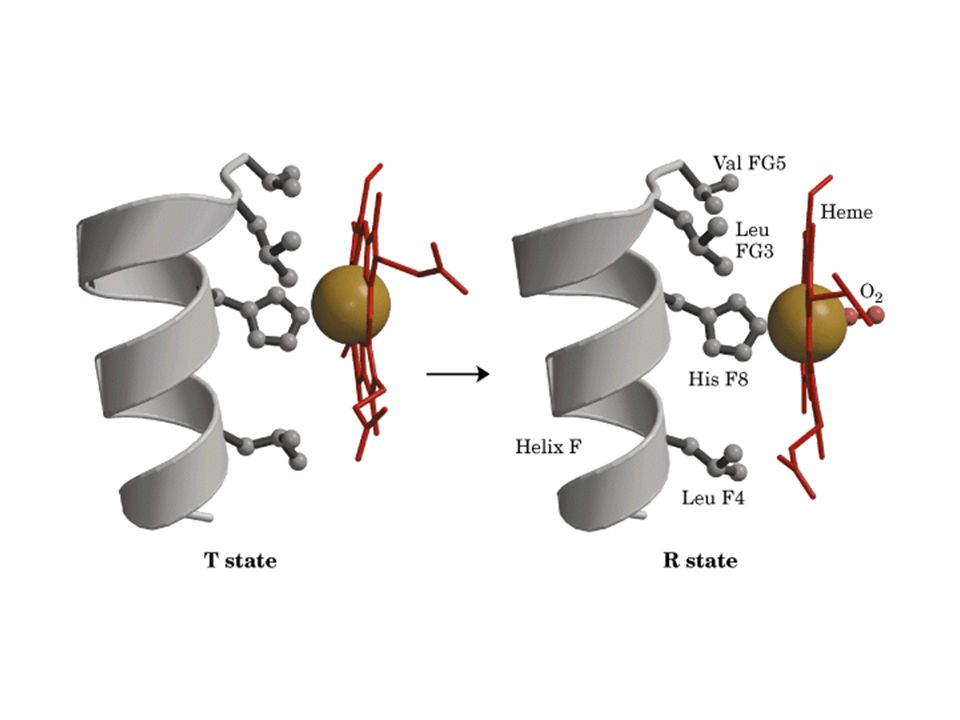
Aminoácidos chaves de mioglobina que interagem com heme e O2
His distal His proximal

66
Conservado em todas as globinas Conservado nas três estruturas

67
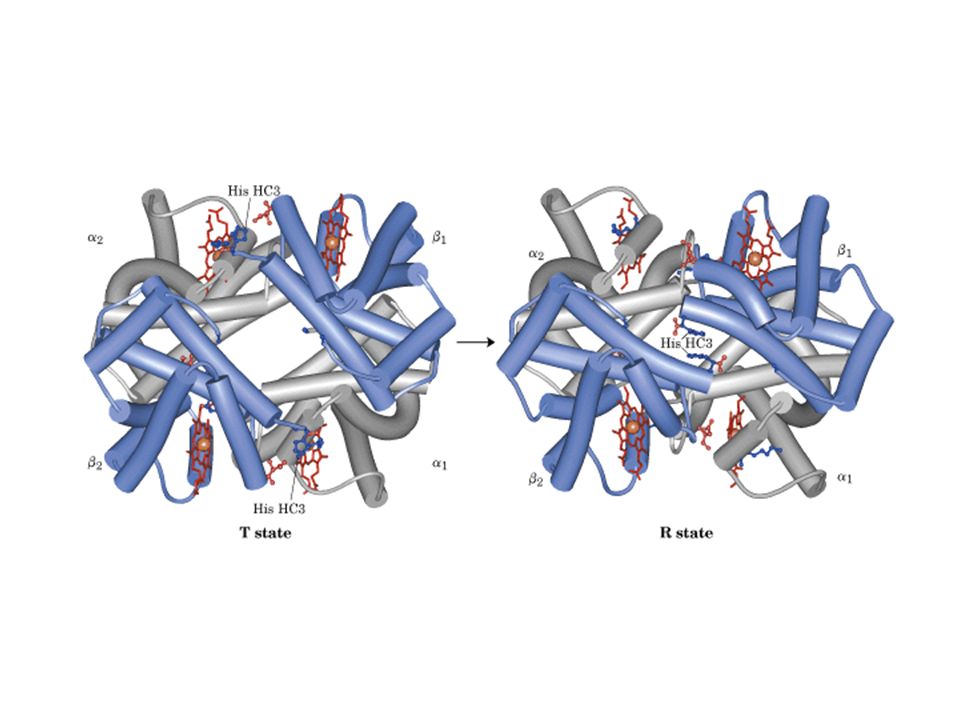
Interações entre subunidades em hemoglobina
Contatos a1-b1 mudam pouco quando O2 liga Contatos a1-b2

68
Contatos a1-b2 no estado T

69
INTERAÇÕES IÔNICAS NO ESTADO T da Hb

72
Ligação de oxigênio a hemoglobina
Curva hiperbólica de Ligação de baixa afinidade Curva sigmoidal de Ligação cooperativa Curva hiperbólica de Ligação de baixa afinidade

73
Dois modelos moleculares para explicar ligação cooperativa
Modelo MWC Monod, Wyman Changeux (“simultânea”) Modelo “sequencial”
 Modelo sequencial")
74
Efeito Bohr (Christian, pai e médico; não Niels, filho e físico)
Hb + O2 HbO2 HHb+ + O2 HbO2 + H+ Logo nos tecidos internos onde [H+] é alto, H+ liga a Hb e causa a dissociação de O2. H+ e O2 ligam em sítios diferentes: -O2 liga no Fe do grupo heme -H+ liga em vários grupos de aminoácidos que estabilizam o estado T (baixa afinidade para O2) ie: His146(+) Asp94(-) Sangue no pulmão Sangue nos tecidos internos
 ie: His146(+) Asp94(-) Sangue. no pulmão. Sangue. nos tecidos. internos.")
75
CO2 + H2N----(a-amino da cadeia)
H+ + -O2C-NH---- terminal “carbamino” Contribui para o efeito Bohr [BPG] alta em hemácias HbO2 + BPG HbBPG + O2

76
HbO2 + BPG HbBPG + O2 No nível de mar, a diferença entre
pO2 nos pulmões e nos tecidos permite o sangue soltar ~40% da sua capacidade de O2 nos tecidos. Em altitudes altas, o forneciemnto de O2 diminuiria para 30% de capacidade sanguina.... Para compensar, a [BPG]aumenta e a afinidade da Hb para O2 cai, resultando no aumento de fornecimeno de O2 para os tecidos até ~40% de sua capacidade.

77
O estado T da Hb tem um sítio para BPG (cargas positivos em azul)
")
78
O estado R da Hb perdeu seu sítio para BPG

79
Hemácias normais

80
Variação de formas de hemácias em anemia falsiforme

81
Anemia falsiforme: uma doença molecular
Glu6 Val na cadeia beta

82
Glu6 Val na cadeia beta resulta num superfície hidrofóbico que promove associação entre moléculas para formar filamentos e fibras

Apresentações semelhantes