Carregar apresentação
A apresentação está carregando. Por favor, espere

1
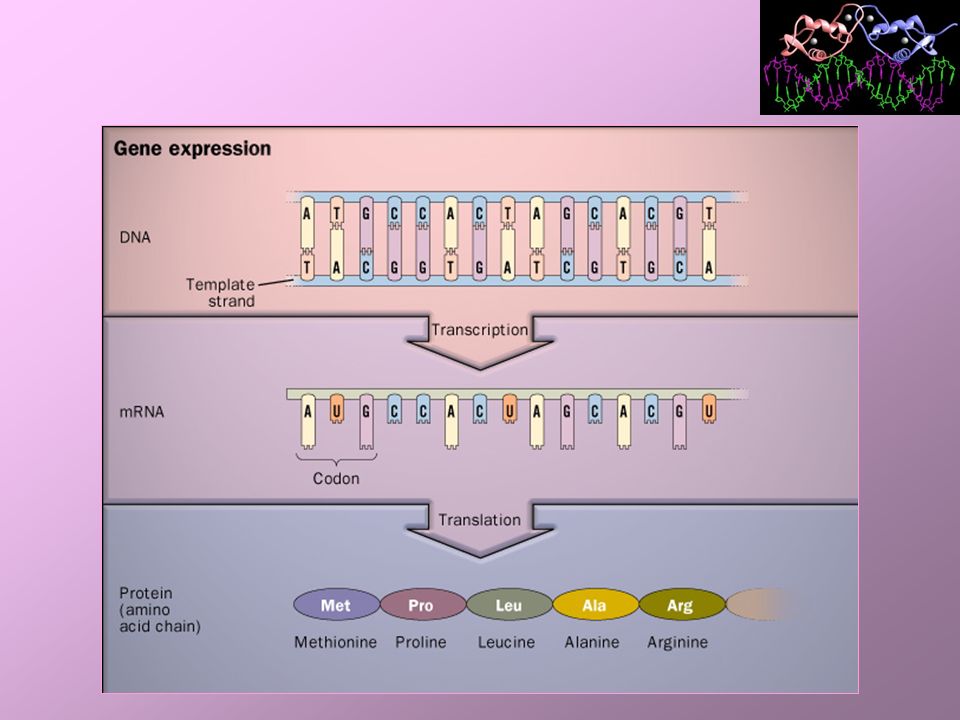
Tradução / Síntese de Proteínas

3
Código Genético “Dicionário” → correspondência da seqüência de nucleotídeos levando à seqüência de aminoácidos; Códon → 3 bases nucleotídicas no RNAm que codificam cada aminoácido (“palavra”); Códon: RNAm → A, G, C e U; “escrita” da direção 5’ para 3’; 64 combinações diferentes de bases;
; Códon: RNAm → A, G, C e U; escrita da direção 5’ para 3’; 64 combinações diferentes de bases;")
4
1 códon 3 nucleotídeos no RNAm
Código Genético 1 códon 3 nucleotídeos no RNAm 7 códons 21 nucleotídeos

5
Código Genético 61 dos 64 códons possíveis codificam os 20 aminoácidos padrão UAG / UGA / UAA Códons de terminação ou de parada ou sem sentido; não codificam AA.

6
Código Genético Características:
Especificidade – um determinado códon sempre codifica o mesmo AA; Universalidade – é conservado em todas as espécies; Redundância ou Degeneração – um AA pode ter mais de 1 trinca que o codifica; Contínuo – sempre lido de 3 em 3 bases. Degeneração do código

7
Mutações no Código Genético
Mutação silenciosa: Códon com 1 base alterada ainda codifica o mesmo AA; Mutação com perda de sentido: Códon com 1 base alterada codifica um AA diferente; Mutação sem sentido: Códon com 1 base alterada se torna um dos códons de terminação;

8
Outras Mutações no Código Genético
Expansão de repetições trinucleotídicas: Inserções de várias repetições de 1 códon. Ex: doença de Huntington; Mutações em sítios de corte-junção: Alteração de íntrons removidos; Mutações com alteração de módulo de leitura: 1 ou 2 nucleotídeos perdidos ou adicionados → seqüência de AAs altera radicalmente.

9
Componentes da Tradução
AAs: Dieta → AAs essenciais; RNAt ou moléculas adaptadoras: Em humanos existem em torno de 50 espécies de RNAt, enquanto bactérias possuem em torno de espécies; Sítios de ligação ao AA – extremidade 3’ do RNAt se liga ao grupo carboxila do AA; Anticódon → seqüência de 3 nucleotídeos que reconhece o códon específico do RNAm; Pode estar carregado ou descarregado.

10
Componentes da Tradução
RNAt: 50 tipos de RNAt para 20 aa: alguns aas possuem mais de um RNAt específico AA é ligado aqui Forma estrutura de folha trevo com 4 braços.Variam em comprimento, de 73 a 93 nucleotídeos. Nucleotídeos extras ocorrem no braço extra ou no braço D. Braço aa e braço anti códon. Braço D e braço TψC O pareamento códon-anticódon é complementar e antiparalelo Estrutura secundária: folha de trevo 1 anticódon pode reconhecer mais de um códon

11
Componentes da Tradução
Aminoacil-RNAt sintetase: Família de enzimas que ligam AA aos seus RNAt → ↑ especificidade que aumenta a fidelidade da tradução da mensagem genética;

12
Componentes da Tradução
RNAm (molde); Ribossomos: Ribossomos: grandes complexos de RNAr e proteínas compostos por duas subunidades. São as estruturas responsáveis pela síntese protéica (local da síntese). Livres ou no RER. Ribossomo de eucariotos: subunidades 60S (5S, 5.8S e 28S/49 proteínas) + 40S (18S/33 proteínas) Em procariotos: subunidades 50S (5S e 23S/36 proteínas) + 30S (16S/21 proteínas) RNAr: responsáveis pela estabilização do complexo de iniciação e dos demais participantes da tradução
; Ribossomos: Ribossomos: grandes complexos de RNAr e proteínas compostos por duas subunidades. São as estruturas responsáveis pela síntese protéica (local da síntese). Livres ou no RER. Ribossomo de eucariotos: subunidades 60S (5S, 5.8S e 28S/49 proteínas) + 40S (18S/33 proteínas) Em procariotos: subunidades 50S (5S e 23S/36 proteínas) + 30S (16S/21 proteínas) RNAr: responsáveis pela estabilização do complexo de iniciação e dos demais participantes da tradução.")
13
Componentes da Tradução
Ribossomos: Sítio P: neste sítio, o códon de iniciação é posicionado para seu pareamento com o anticódon do RNAt que transposta metionina – primeiro aa da tradução. Sítio A: neste sítio, o códon adjacente é posicionado para seu pareamento com o anticódon do RNAt que transposta o próximo aa da cadeia polipeptídica. Sítio E: depois de ser traduzido, o códon é posicionado no sítio E (ou sítio de saída) para seu desligamento com o RNAt, agora descarregado. Fatores protéicos: Fatores de iniciação, alongamento e terminação ou liberação. ATP e GTP.
 para seu desligamento com o RNAt, agora descarregado. Fatores protéicos: Fatores de iniciação, alongamento e terminação ou liberação. ATP e GTP.")
14
Reconhecimento dos Códons pelo RNAt
A ligação entre códon do RNAm e anticódon do RNAt é antiparalela; O códon é lido de 5’ para 3’; o anticódon também deve ser lido de 5’ para 3’. Portanto a primeira base do códon pareia com a última base do anticódon;

15
Reconhecimento dos Códons pelo RNAt
Hipótese da Oscilação: Se a trinca do anticódon reconhecesse apenas 1 trinca do códon por pareamento, as células deveriam ter 1 RNAt para cada códon de AA → NÃO É O QUE OCORRE!; As 2 primeiras bases do códon formam pares de bases Watson-Crick com fortes pontes de hidrogênio → dão especificidade da codificação; A terceira base do códon que pareia com a primeira base do anticódon forma pontes de hidrogênio mais fracas e a primeira base do anticódon pode parear com mais de 1 base. Para alguns aa diferentes codóns codificam para o mesmo aa. Como por exemplo a arginina que quatro códons codificam para este aa. Mas quanto ao anti-códon, Há quatro RNAt, um para cada codificar cada códon? Não. Porque? Porque…As duas primeiras bases do códon são idênticas (CG) formando fortes pares de base Watson-Crick com as bases correspondentes do antí-códon, porém a terceira base da maioria dos códons pareia de maneira frouxa com a base correspondente de seus anticódons, ou seja, as terceiras bases de tais códons (e as primeiras de seus anticódons correspondentes) oscilam.
 formando fortes pares de base Watson-Crick com as bases correspondentes do antí-códon, porém a terceira base da maioria dos códons pareia de maneira frouxa com a base correspondente de seus anticódons, ou seja, as terceiras bases de tais códons (e as primeiras de seus anticódons correspondentes) oscilam.")
16
Hipótese da Oscilação 1 anticódon pareia com mais de 1 códon!
Lig. + específica Lig. - específica A base oscilante (ou terceira) do códon contribui para a especificidade, mas , pelo fato dela parear apenas fracamente com sua base correspondente no anticódon, ela permite rápida dissociação do tRNA de seu códon, durante a síntese de proteína. Se todas as três bases dos códons estivessem engajadas em pareamentos Watson-cricK fortes com as três bases dos anticódons, os tRNAs se dissociariam muito lentamente, o que limitaria severamente a velocidade da síntese de proteínas. As interações condón-anticódon otimizam tanto a exatidão quanto a velocidade. OBS: I (inosina) contém base hipoxantina pode ser encontrado como a primeira base do anticódon → base oscilante que pareia com mais de 1 base As interações códon-anticódon otimizam tanto a exatidão quanto a velocidade de síntese protéica
 do códon contribui para a especificidade, mas , pelo fato dela parear apenas fracamente com sua base correspondente no anticódon, ela permite rápida dissociação do tRNA de seu códon, durante a síntese de proteína. Se todas as três bases dos códons estivessem engajadas em pareamentos Watson-cricK fortes com as três bases dos anticódons, os tRNAs se dissociariam muito lentamente, o que limitaria severamente a velocidade da síntese de proteínas. As interações condón-anticódon otimizam tanto a exatidão quanto a velocidade. OBS: I (inosina) contém base hipoxantina pode ser encontrado como a primeira base do anticódon → base oscilante que pareia com mais de 1 base. As interações códon-anticódon otimizam tanto a exatidão quanto a velocidade de síntese protéica.")
17
Etapas da Síntese Protéica
Ativação dos AAs: Ligação dos AAs aos seus RNAt ocorre no citosol pelas aminoacil-RNAt sintetases. Duas lig. de alta energia Aminoacilação do RNAt No citosol os aa são esterificados aos seus correspondentes RNAt através da enzima aminoacil-RNAt sintetase. Cada enzima é específica para um aa e uma ou mais RNAt correspondentes. A reação de ativação ocorre em duas etapas no sítio ativo da enzima eforma-se a lig. éster entre o aa e o RNAt. Então ocorreu a aminoacilação do RNAt que realiza a ativação do aa para a formação da lig. Polipeptídica e a lig. do aa a um RNAt adaptador, que garante sua colocação apropriada dentro de um polipeptídeo em crescimento. Requer: 20 aas 20 aminoacil-tRNA sintetases Energia – ATP RNAt

18
Etapas da Síntese Protéica
Iniciação: O RNAm liga-se a menor das 2 subunidades ribossômicas e ao aminoacil-RNAt de iniciação; Na E. coli, a seqüência reconhecida no RNAm pelo ribossomo é chamada de seqüência de Shine-Dalgarno (nos eucariotos o “quepe” do RNAm é reconhecido pelo ribossomo) → 6 a 10 bases longe do códon de iniciação AUG;
 → 6 a 10 bases longe do códon de iniciação AUG;")
19
Etapas da Síntese Protéica
Iniciação: O aminoacil-RNAt de iniciação pareia com o códon AUG, que é o códon que sinaliza o início da proteína a ser sintetizada; Em bactérias e na mitocôndria, esse RNAt de iniciação carrega uma metionina N-formilada (grupo formila é adicionado pela enzima transformilase). Nos eucariotos, a metionina não está formilada; A síntes e de proteínas começa na extremidade aminoterminal e prossegue pela adição passo a passo dos aas. O códon de inicaçãoAUG especifica um resíduo de metionina aminoterminal. A pesar da metionina apresentar somente um códon 5´AUG, todos os organismos apresentam dois tRNA para metionina. Um é usado exclusivamente quando AUG é o códon de iniciação para a síntese de protínas. O segundo é usado para codificar metionina em uma posição interna em um polipeptídeo. Um é formilado (posição inicial) e o outro (interna) não
. Nos eucariotos, a metionina não está formilada; A síntes e de proteínas começa na extremidade aminoterminal e prossegue pela adição passo a passo dos aas. O códon de inicaçãoAUG especifica um resíduo de metionina aminoterminal. A pesar da metionina apresentar somente um códon 5´AUG, todos os organismos apresentam dois tRNA para metionina. Um é usado exclusivamente quando AUG é o códon de iniciação para a síntese de protínas. O segundo é usado para codificar metionina em uma posição interna em um polipeptídeo. Um é formilado (posição inicial) e o outro (interna) não.")
20
Etapa 2 Iniciação Cofator enzimático – Mg+2 Requer: RNAm
Requer a subunidade ribossômica 30S, RNAm que codifica o polipeptídeo a ser sintetizado, fMe-RNAt de iniciação e um conjunto de 3 proteínas, chamados de fatores de iniciação e Mg2+. A formação do complexo de iniciação realiza-se em 3 etapas: Etapa 1: A sequencia Shine – Dalgarno guia a posição correta no RNAm da localização do códon de iniciação 5´AUG Requer: RNAm aminoacil-tRNA de iniciação – metionina códon de iniciação - AUG Subunidade 30S e 50S Fatores de iniciação GTP Cofator enzimático – Mg+2

21
Formação do complexo de iniciação em eucariotos
A tradução eucariota é geralmente semelhante a procariota, com a maioria da diferenças significativas no mecanismode iniciação. Vários fatores de iniciação estão envolvidos no processo de iniciação da síntese protéica nos eucariotos (complexo eIF4F) diferindo um pouco dos procariotos.Acredita-se que elas mantenham juntas as extremidades 5´e 3´da mensagem. Nos eucariotos o “quepe” do RNAm é reconhecido pelo ribossomo. O complexo proteico eIF4F liga-se tanto a região cap por meio de eIF4E quento a região poli A por meio de PAB.
 diferindo um pouco dos procariotos.Acredita-se que elas mantenham juntas as extremidades 5´e 3´da mensagem. Nos eucariotos o quepe do RNAm é reconhecido pelo ribossomo. O complexo proteico eIF4F liga-se tanto a região cap por meio de eIF4E quento a região poli A por meio de PAB.")
22
Etapas da Síntese Protéica
Alongamento: Fatores de alongamento são necessários (EF-Tu, EF-Ts, EF-G); Peptidiltransferase (ribozima) → liga o peptídeo em formação e o AA a ser adicionado; Após a ligação peptídica se formar, o ribossomo avança 3 nucleotídeos na direção 3’→Translocação (requer energia, GTP) . O RNAt não-carregado vai para o sítio E antes de ser liberado e o RNAt carregando o peptídeo vai para o sítio P .
; Peptidiltransferase (ribozima) → liga o peptídeo em formação e o AA a ser adicionado; Após a ligação peptídica se formar, o ribossomo avança 3 nucleotídeos na direção 3’→Translocação (requer energia, GTP) . O RNAt não-carregado vai para o sítio E antes de ser liberado e o RNAt carregando o peptídeo vai para o sítio P .")
23
Etapa 3 Alongamento Requer: Complexo de iniciação aminoacil-tRNA
especificados pelos códons Fatores de alongamento Peptidiltransferase GTP Vamos ver melhor isso.

24
Alongamento Tanslocação
Etapa 3 O ribossomo se move em direção à extremidade 3´do mRNA, o peptidil-tRNA está, agora, no sítio P deixando o sítio A aberto para o terceiro aminoacil-tRNA. O tRNA não-carregado é deslocado para o sítio E, desligando-se imediatamente do ribossomo. A translocação envolve o complexo fator de elongação EF-G-GTP.

25
Etapa 4 Terminação: Ocorre quando 1 dos 3 códons (UAA, UAG, UGA) de terminação é “colocado” no sítio A; Na E. coli, os fatores de terminação ou liberação reconhecem esses códons e ocorre a liberação do complexo ribossomal. O alongamento continua até que o ribossomo adicione o último aa codificado pelo RNAm

26
Etapa 4 Terminação -Hidrólise da lig. peptidil-RNAt terminal;
-Liberação do peptídeo livre e do RNAt; -Dissociação do ribossomo 70S. Nas bactérias, assim que um códon de terminação ocupa o sítio ribossomico A, três fatores de liberação ou terminação, as proteínas RF1, RF2, RF3 contribuem para … O RF1 reconhece os códons de terminação UAG e UAA, e o RF2 reconhece UGA e UAA. Ou o RF1 ou o RF2 (dependendo de que códon esteja presente) liga-se ao códon de terminação e induz a peptidiltransferase a transferir a cadeia polipeptídica crescente para uma molécula de água em vez de para um outro aa. Requer: Códons de terminação Fatores de liberação
 liga-se ao códon de terminação e induz a peptidiltransferase a transferir a cadeia polipeptídica crescente para uma molécula de água em vez de para um outro aa. Requer: Códons de terminação. Fatores de liberação.")
27
Etapas da Síntese Protéica
Polissomos ou Polirribossomos: Complexo de 1 RNAm e vários ribossomos. Grandes agregados de 10 a 100 ribossomos que são muito ativos na síntese proteéica podem ser isolados tanto de células eucariotas quanto de bacterianas. A fita conectante é uma única fita de RNAm, sendo traduzida simultaneamente por muito ribossomos cuidadosamente espaçados, permitindo o uso altamente eficiente do RNAm. Direção da transcrição e posicionamento dos ribossomos 5´, 3´ Nas bactérias a transcrição e tradução estão fortemente acopladas porque não apresentam a membrana nuclear. A fim de manter altas taxas de síntese de proteínas, o RNAm de uma certa proteínas, ou conjunto de proteínas, deve ser sintetizado continuamente e traduzido com a máxima eficiência.

28
Modificações Pós-translacionais e a Estrutura Tridimensional
Etapa 5: Modificações Pós-translacionais e a Estrutura Tridimensional Após a tradução, algumas proteínas, antes de assumirem a sua conformação nativa, têm a sua estrutura primária alterada por modificações pós-translacionais, como por exemplo: Fosforilação Carboxilação Na quinta e última etapa da síntese de proteínas, a cadeia polipeptídica nascente é enrolada e processada na sua forma biologicamente ativa.Durante e após a síntese o polipeptídeo assume progressivamente a sua conformação nativa, com a formação apropriada de pontes de hidrogênio e das interações de vander Waaals, iônicas e hidrofóbicas. Desta forma, a mensagem linear ou unidimensional do RNAm é convertida na estrutura tridimencional da proteína. Algumas proteínas recém-sintetizadas, tanto procariota quanto eucariota, não atingem sua conformação final biologicamente ativa, até que tenham sido alteradas por uma ou mais reações de processamento, chamadas de m odificações pós-traducionais. Inicialmente, todos os polipeptídeos começam com um resíduo N-formilmetionina (na bactéria) ou metionina (nos eucariotos). Entretanto o grupo formila, s resíduo aminoterminal Met e frequentemente outros resíduos amino terminais podem ser removidos enzimaticamente e não aparecem nas proteínas funcionais finais. Sequencia sinalizadora (15 a 30 aa) para direcioanmento da proteína são removidas no final. Exemplo da caseína, proteína do leite, possue muito grupos de fosfoserina. Grupo fostato adiciona carga negativa. Desta forma liga-se calcio. Ca, fosfato e aa são impotantes nutrientes para o lactente. Prototrombina- vários resíduos de γ-carboxilglutamato em sua região aminoterminal e estes se ligam ao Ca requerido para iniciar o processo de coagulação Protrombina
 ou metionina (nos eucariotos). Entretanto o grupo formila, s resíduo aminoterminal Met e frequentemente outros resíduos amino terminais podem ser removidos enzimaticamente e não aparecem nas proteínas funcionais finais. Sequencia sinalizadora (15 a 30 aa) para direcioanmento da proteína são removidas no final. Exemplo da caseína, proteína do leite, possue muito grupos de fosfoserina. Grupo fostato adiciona carga negativa. Desta forma liga-se calcio. Ca, fosfato e aa são impotantes nutrientes para o lactente. Prototrombina- vários resíduos de γ-carboxilglutamato em sua região aminoterminal e estes se ligam ao Ca requerido para iniciar o processo de coagulação. Protrombina.")
29
Modificações Pós-translacionais e a Estrutura Tridimensional
Etapa 5: Modificações Pós-translacionais e a Estrutura Tridimensional Metilação Outras Modificações Monometil e dimetilisina-Proteínas musculares e citocromoc Trimetilisina- Calmodulina

30
Modificações Pós-translacionais e a Estrutura Tridimensional
Etapa 5: Modificações Pós-translacionais e a Estrutura Tridimensional As proteínas assumem a sua conformação nativa com o auxílio das chaperonas ou proteínas do estresse ou proteínas do choque térmico (heat shock proteins). Ajudam as proteínas a se moldar, associar a outras proteínas de maneira estável e tornarem-se estruturas ativas, evitando a associação de proteínas ainda não dobradas corretamente Cada Chaperona possui um compartimento central, que acomoda a proteína recém-sintetizada, impedindo que esta tenha um dobramento prematuro e de forma inadequada. Assim, as Chaperonas ajudam as proteínas a se moldar, associar a outras proteínas de maneira estável e tornarem-se estruturas ativas, evitando a associação de proteínas ainda não dobradas corretamente. Também atuam ativamente, com hidrólise de ATP e modificação conformacional de algumas proteínas. Conformação Desnaturada Conformação Nativa
. Ajudam as proteínas a se moldar, associar a outras proteínas de maneira estável e tornarem-se estruturas ativas, evitando a associação de proteínas ainda não dobradas corretamente. Cada Chaperona possui um compartimento central, que acomoda a proteína recém-sintetizada, impedindo que esta tenha um dobramento prematuro e de forma inadequada. Assim, as Chaperonas ajudam as proteínas a se moldar, associar a outras proteínas de maneira estável e tornarem-se estruturas ativas, evitando a associação de proteínas ainda não dobradas corretamente. Também atuam ativamente, com hidrólise de ATP e modificação conformacional de algumas proteínas. Conformação. Desnaturada. Conformação. Nativa.")
31
Diferenças entre procariotos e eucariotos
Ligação do mRNA à subunidade menor ribossomal O 5´-CAP do mRNA liga-se aos fatores de iniciação e à subunidade 40S. O mRNA é lido a partir do códon de iniciação Primeiro Aminoácido Metionina (não formilada) Fatores de iniciação eIFs (8 ou mais) Fatores de terminação eRF Fatores de alongamento EF1 a ( EF-Tu) EF bg ( EF-Ts) EF2 (EF-G) Ribossomo 80S (40S + 60S) ausência de sítio E ( exit tradução não simultânea com transcrição
 Fatores de iniciação. eIFs (8 ou mais) Fatores de terminação. eRF. Fatores de alongamento. EF1. a ( EF-Tu) EF. bg. ( EF-Ts) EF2 (EF-G) Ribossomo. 80S (40S + 60S) ausência de sítio E ( exit. tradução não simultânea com. transcrição.")
32
Diferenças e semelhanças entre eucariotos e procariotos
O esquema geral é o mesmo, e a síntese em si ocorre em estruturas similares: os ribossomos Mas, em eucariotos a transcrição está separada da síntese de proteínas (tradução) pela membrana nuclear
 pela membrana nuclear.")
33
Retículo Endoplasmático Rugoso
Síntese Protéica no Retículo Endoplasmático Rugoso Síntese protéica no retículo endoplasmático rugoso daquelas proteínas que serão localizadas na membrana plasmática ou nos lisossomos, ou serão secretadas. Proteínas destinadas à secreção, integração na membrna plasmática, ou inclusão nos lisossomos, geralmente compartilham algumas etapas iniciais de uma via que começa no RE. Proteínas destinadas às mitocôndrias e núcleo usam très mecanismos separados. Proteínas destinadas ao citosol simplesmente permanecem onde foram sintetizadas. Estas proteínas apresentam sequencias sinalizadoras que se localizam na região aminoterminal de um peptídeo recém-sintetizado. Esta sequencia é removida durante o transporte para o local final ou no local final. A via de endereçamento começa com a iniciação da síntese de proteínas nos ribossomos livres. A sequência sinalizadora aparece no começo do processo sintetizante, porque ela está no aminoterminal, que é sintetizado primeiro. À medida que ela emerge do ribossomo, essa sequência e o próprio ribossomo são rapidamente ligados pela grande partícula de reconhecimento da sinalização (SRP). Esta ligada ao GTP direciona o ribossomo (ainda ligado ao RNAm) e o polipeptídeo incompleto para receptores da SRP na face citosólica do RE. O polipeptídeo nascente é entregue a um complexo de translocação do peptídeo no RE, que pode interagir diretamente com o ribossomo. SRP dissocia-se do ribossomo.O alongamento do polipeptídeo recomeça, até que a proteína completa tenha sido sintetizada. A sequencia sinalizadora é removida por um peptidase dentro da luz do RE. O ribossomo dissocia e é reciclado.
. Esta ligada ao GTP direciona o ribossomo (ainda ligado ao RNAm) e o polipeptídeo incompleto para receptores da SRP na face citosólica do RE. O polipeptídeo nascente é entregue a um complexo de translocação do peptídeo no RE, que pode interagir diretamente com o ribossomo. SRP dissocia-se do ribossomo.O alongamento do polipeptídeo recomeça, até que a proteína completa tenha sido sintetizada. A sequencia sinalizadora é removida por um peptidase dentro da luz do RE. O ribossomo dissocia e é reciclado.")
34
Síntese Protéica no RER
Síntese proteíca no reticulo endoplasmático rugoso daquelas proteínas que serão localizadas na membrana plasmática ou nos lisossomos, ou serão secretadas Síntese Protéica no RER Aquí uma visão geral deste processo. Ribossomos associados ao MRE sintetizando proteínas para a luz do RE (que apresentam a região sinalizadora que enderam para compor MP, lisossomos ou serem secretadas)
")
35
Inibidores de Síntese Protéica
Estreptomicina: Liga-se à subunidade 30S e distorce sua estrutura inibindo a iniciação Tetraciclina: bloqueia o sítio A ribossomal X X

36
Inibidores de Síntese Protéica
Cloranfenicol: Inibe a atividade de peptidil- transferase procariótica Clindamicina e Eritromicina: Ligam-se de maneira irreversível à subunidade 50S do ribossomo bacteriano, inibindo o deslocamento. X X

37
Bibliografia: Voet, D., Voet, J.G., Pratt, C.W. Fundamentos de Bioquímica (2000). Champe, P.C.; Harvey, R.A.; Ferrier, D.R. Bioquímica Ilustrada (3a ed, 2006). Nelson, D.L., Cox, M.M. Lehninger, Princípios de Bioquímica. Quarta edição (2004).
. Nelson, D.L., Cox, M.M. Lehninger, Princípios de Bioquímica. Quarta edição (2004).")
Apresentações semelhantes