Carregar apresentação
A apresentação está carregando. Por favor, espere

1
FIGURA 6. 2Genoma do vírus símio 40 (SV40)
FIGURA 6.2Genoma do vírus símio 40 (SV40). DNA de SV40, apresentado em tracejado preto, é um círculo de fita dupla de pouco mais que 5000 pb que codifica toda a infor-mação necessária à sobrevivência e replica-ção do vírus dentro de uma célula hospedei-ra. Demonstra uso extremamente eficiente do potencial de codificação de proteína de um genoma. Proteínas VP1, VP2 e VP3 são proteínas estruturais do vírus; VP2 e VP3 são traduzidas a partir de diferentes pontos de iniciação até a mesma extremidade carboxi-terminal. VP1 é traduzida em uma fase de leitura diferente, de modo que sua porção N-terminal se sobrepõe aos genes VP2 e VP3, mas sua seqüência de aminoácidos no segmento sobreposto é diferente de VP2 e VP3. Duas proteínas, os antígenos tumorais large T (T grande) e small t (t pequeno), que promovem transformação de células infec-tadas, são codificadas por um precursor de mRNA comum. Ambas as proteínas têm se-qüências N-terminais idênticas; o segmento C-terminal da proteína small t é codificado por um segmento de mRNA, que é remo-vido da mensagem da proteína large T, e a seqüência carboxi-terminal de large T é co-dificada no RNA, que se segue à terminação de small t. A origem da replicação do DNA (ori) fica fora das regiões codificadoras.
. DNA de SV40, apresentado em tracejado preto, é um círculo de fita dupla de pouco mais que 5000 pb que codifica toda a infor-mação necessária à sobrevivência e replica-ção do vírus dentro de uma célula hospedei-ra. Demonstra uso extremamente eficiente do potencial de codificação de proteína de um genoma. Proteínas VP1, VP2 e VP3 são proteínas estruturais do vírus; VP2 e VP3 são traduzidas a partir de diferentes pontos de iniciação até a mesma extremidade carboxi-terminal. VP1 é traduzida em uma fase de leitura diferente, de modo que sua porção N-terminal se sobrepõe aos genes VP2 e VP3, mas sua seqüência de aminoácidos no segmento sobreposto é diferente de VP2 e VP3. Duas proteínas, os antígenos tumorais large T (T grande) e small t (t pequeno), que promovem transformação de células infec-tadas, são codificadas por um precursor de mRNA comum. Ambas as proteínas têm se-qüências N-terminais idênticas; o segmento C-terminal da proteína small t é codificado por um segmento de mRNA, que é remo-vido da mensagem da proteína large T, e a seqüência carboxi-terminal de large T é co-dificada no RNA, que se segue à terminação de small t. A origem da replicação do DNA (ori) fica fora das regiões codificadoras.")
2
FIGURA 6. 3Interação de um tRNA com sua aminoacil-tRNA sintetase
FIGURA 6.3Interação de um tRNA com sua aminoacil-tRNA sintetase. O esqueleto açúcar-fosfato do glutaminil tRNA de E. coli é apresentado em cinza-médio, e o esqueleto peptídico da glutaminil-tRNA sintetase é apresentado em múltiplos tons. Note as interações da sintetase com haste aceptora parcialmente desenrolada e alça do anticódon do tRNA; note também a localização do ATP, mostrado em cinza-escuro dentro do retângulo, a poucos Angstroms da extremidade 3’ do tRNA. Modelos de preenchimento de espaço da enzima e do tRNA mostram ambas as moléculas como objetos sólidos, com vários pontos de contato direto.Adaptado de Perona, J., Rould, M. e Steitz, T. Biochemistry 32:8758, 1993.

4
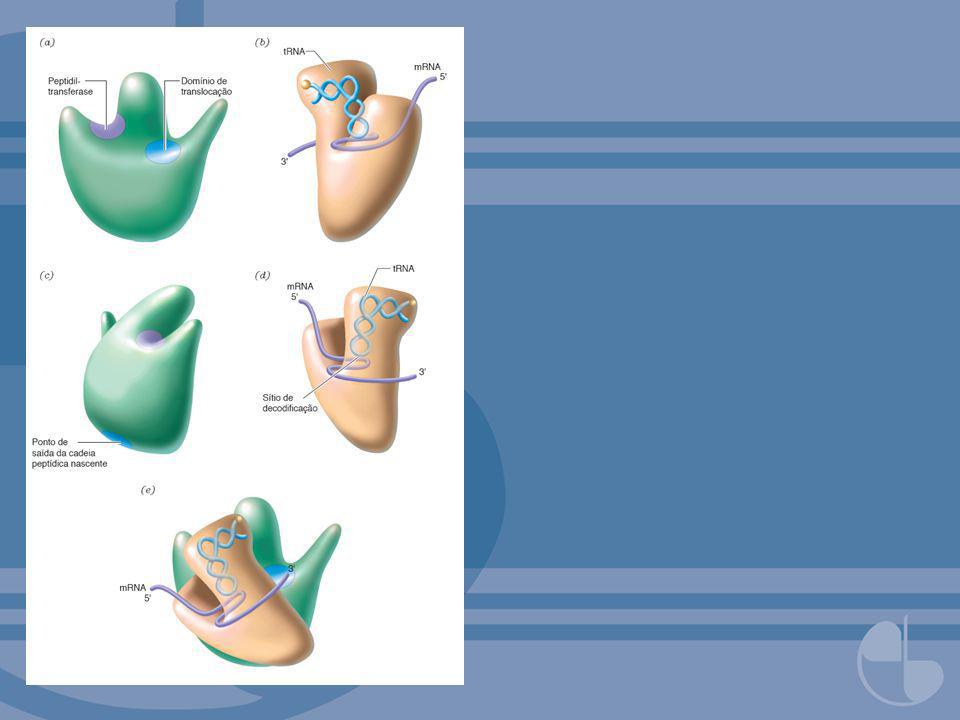
FIGURA 6. 5Estrutura cristalográfica de um ribossomo 70 S
FIGURA 6.5Estrutura cristalográfica de um ribossomo 70 S. A estrutura do ribossomo de Thermus thermophilus é mostrada a uma resolução de 5,5 Å. Vistas sucessivas são rotações de 90º em torno do eixo vertical. (a) A subunidade pequena está sobre a subunidade grande, como no modelo da Figura 6.4e. Características da subunidade pequena incluem a plataforma (P), a cabeça (H), o corpo (B) e um pescoço (N) que faz a conexão. O 16 S RNA está em cinza-claro, e proteínas da subunidade pequena estão em cinza-escuro. (b) A subunidade grande está à direita; 23 S RNA é cinza, 5 S RNA é cinza mais claro, e proteínas da subunidade grande são púrpuras. Uma molécula de tRNA no sítio A (cinza) atravessa as subunidades. (c) A subunidade grande está em cima, com sua haste se projetando à esquerda. (d) A subunidade grande está à esquerda e elementos de tRNA são visíveis na interface das subunidades.Reimpresso com permissão de Yusupov, M. M., Yusupova, G. Z., Baucom, A., Lieberman, K., Earnest, T. N., Cate, J. H. D. e Noller, H. F. Science 292:883, Direitos autorais (2001) AAAS. Figura gentilmente cedida por Drs. A. Baucom e H. Noller.
 A subunidade pequena está sobre a subunidade grande, como no modelo da Figura 6.4e. Características da subunidade pequena incluem a plataforma (P), a cabeça (H), o corpo (B) e um pescoço (N) que faz a conexão. O 16 S RNA está em cinza-claro, e proteínas da subunidade pequena estão em cinza-escuro. (b) A subunidade grande está à direita; 23 S RNA é cinza, 5 S RNA é cinza mais claro, e proteínas da subunidade grande são púrpuras. Uma molécula de tRNA no sítio A (cinza) atravessa as subunidades. (c) A subunidade grande está em cima, com sua haste se projetando à esquerda. (d) A subunidade grande está à esquerda e elementos de tRNA são visíveis na interface das subunidades.Reimpresso com permissão de Yusupov, M. M., Yusupova, G. Z., Baucom, A., Lieberman, K., Earnest, T. N., Cate, J. H. D. e Noller, H. F. Science 292:883, Direitos autorais (2001) AAAS. Figura gentilmente cedida por Drs. A. Baucom e H. Noller.")
5
FIGURA 6. 6Micrografias eletrônicas de polissomos
FIGURA 6.6Micrografias eletrônicas de polissomos. (a) Polirribossomos de reticulócitos sombreados com platina são vistos em grupos de três a seis ribossomos, um número consistente com o tamanho do mRNA de uma cadeia de globina. (b) Coloração com uranil acetato e visualização em maior aumento mostram polissomos, nos quais partes do mRNA são visíveis.Cortesia de Dr. Alex Rich, MIT.
 Polirribossomos de reticulócitos sombreados com platina são vistos em grupos de três a seis ribossomos, um número consistente com o tamanho do mRNA de uma cadeia de globina. (b) Coloração com uranil acetato e visualização em maior aumento mostram polissomos, nos quais partes do mRNA são visíveis.Cortesia de Dr. Alex Rich, MIT.")
6
FIGURA 6. 7Iniciação da tradução em eucariotos. Detalhes no texto
FIGURA 6.7Iniciação da tradução em eucariotos. Detalhes no texto. Um complexo ternário que inclui o tRNA iniciador (etapa 1) combina-se com uma subunidade ribossômica pequena (etapa 2). Interação com mRNA forma um complexo de pré-iniciação (etapa 3). Ligação da subunidade grande completa formação do complexo de iniciação (etapa 4). A forma diferente de eIF2a complexado com GTP ou GDP indica que mudanças conformacionais ocorrem na proteína com hidrólise do trifosfato. Depois do início da elongação, subunidades pequenas adicionais se complexarão com o mesmo mRNA para formar polissomos.
 combina-se com uma subunidade ribossômica pequena (etapa 2). Interação com mRNA forma um complexo de pré-iniciação (etapa 3). Ligação da subunidade grande completa formação do complexo de iniciação (etapa 4). A forma diferente de eIF2a complexado com GTP ou GDP indica que mudanças conformacionais ocorrem na proteína com hidrólise do trifosfato. Depois do início da elongação, subunidades pequenas adicionais se complexarão com o mesmo mRNA para formar polissomos.")
7
FIGURA 6. 8Etapas de elongação em síntese protéica eucariótica
FIGURA 6.8Etapas de elongação em síntese protéica eucariótica. (a) O primeiro ciclo de elongação é mostrado. Etapa 1: complexo de iniciação com metionil tRNAimet no sítio 80 S P. Etapa 2: EF1α colocou um aminoacil-tRNA no sítio A. Hidrólise de GTP resulta em mudança de conformação de EF1α. Etapa 3: a primeira ligação peptídica se formou, o novo peptidil-tRNA ocupa um sítio híbrido (A/P) no ribossomo e o braço aceptor desacilado do tRNAimet está no sítio E da subunidade grande. Etapa 4: o complexo mRNA-peptidil-tRNA foi translocado para o sítio P, enquanto o tRNA iniciador desacilado move-se para o sítio E. (b) Ciclos subseqüentes de elongação. Etapa 5: ligação do aminoacil-tRNA no sítio A causa liberação do tRNA desacilado do sítio E. Etapa 6: formação da ligação peptídica resulta no novo peptidil-tRNA ocupando um sítio híbrido A/P do ribossomo. Etapa 7: translocação move mRNA e novo peptidil-tRNA, em registro, para sítio P. Aminoácidos adicionais são colocados por repetições sucessivas do ciclo. Para maiores detalhes, ver texto.
 O primeiro ciclo de elongação é mostrado. Etapa 1: complexo de iniciação com metionil tRNAimet no sítio 80 S P. Etapa 2: EF1α colocou um aminoacil-tRNA no sítio A. Hidrólise de GTP resulta em mudança de conformação de EF1α. Etapa 3: a primeira ligação peptídica se formou, o novo peptidil-tRNA ocupa um sítio híbrido (A/P) no ribossomo e o braço aceptor desacilado do tRNAimet está no sítio E da subunidade grande. Etapa 4: o complexo mRNA-peptidil-tRNA foi translocado para o sítio P, enquanto o tRNA iniciador desacilado move-se para o sítio E. (b) Ciclos subseqüentes de elongação. Etapa 5: ligação do aminoacil-tRNA no sítio A causa liberação do tRNA desacilado do sítio E. Etapa 6: formação da ligação peptídica resulta no novo peptidil-tRNA ocupando um sítio híbrido A/P do ribossomo. Etapa 7: translocação move mRNA e novo peptidil-tRNA, em registro, para sítio P. Aminoácidos adicionais são colocados por repetições sucessivas do ciclo. Para maiores detalhes, ver texto.")
8
FIGURA 6.8(continuação)
")
9
FIGURA 6. 9EF1 no ciclo de elongação
FIGURA 6.9EF1 no ciclo de elongação. Etapa 1: complexo EF1α-GTP-aminoacil-tRNA liga-se ao ribossomo. Etapa 2: aminoacil-tRNA é colocado no ribossomo (2a), com hidrólise de GTP e uma mudança de conformação de EF1α (Etapa 3), que reduz sua afinidade por tRNA e ribossomo. Etapa 4: GDP é deslocado de EF1α por EF1βγ. Ligação de GTP, então, desloca EF-1βγ (Etapa 5) e permite ligação de um aminoacil-tRNA a EF1α em sua conformação de maior afinidade (1).
, com hidrólise de GTP e uma mudança de conformação de EF1α (Etapa 3), que reduz sua afinidade por tRNA e ribossomo. Etapa 4: GDP é deslocado de EF1α por EF1βγ. Ligação de GTP, então, desloca EF-1βγ (Etapa 5) e permite ligação de um aminoacil-tRNA a EF1α em sua conformação de maior afinidade (1).")
10
FIGURA 6. 10Terminação da síntese protéica
FIGURA 6.10Terminação da síntese protéica. Quando um códon de terminação no mRNA ocupa o sítio A, ligação de um complexo fator de liberação-GTP ocorre (Etapa 1) e tRNA desacilado é liberado. Etapa 2: peptidil transferase funciona como hidrolase; hidrólise da ligação éster que a proteína ao tRNA libera a proteína. A extremidade aceptora do tRNA desacilado é provavelmente deslocada. GTP é hidrolisado, e fator de liberação-GDP dissocia. Componentes dissociados podem agora entrar em ciclos adicionais de síntese protéica.
 e tRNA desacilado é liberado. Etapa 2: peptidil transferase funciona como hidrolase; hidrólise da ligação éster que a proteína ao tRNA libera a proteína. A extremidade aceptora do tRNA desacilado é provavelmente deslocada. GTP é hidrolisado, e fator de liberação-GDP dissocia. Componentes dissociados podem agora entrar em ciclos adicionais de síntese protéica.")
11
FIGURA 6. 12Retículo endoplasmático rugoso
FIGURA 6.12Retículo endoplasmático rugoso. Três setas paralelas indicam três ribossomos dentre os muitos ligados às membranas. Seta única indica uma mitocôndria, para comparação.Cortesia de Dr. U. Jarlfors, University of Miami.

12
FIGURA 6. 13Via secretória: reconhecimento do peptídeo sinal
FIGURA 6.13Via secretória: reconhecimento do peptídeo sinal. Etapa A: um peptídeo sinal hidrofóbico emerge de um ribossomo livre para o citosol. Etapa B: partícula de reconhecimento de sinal (SRP) liga-se ao peptídeo sinal e elongação é temporariamente paralisada. Etapa C: o ribossomo move-se para a membrana do ER, onde uma proteína de aportamento (docking) liga-se a SRP. Etapa D: ribossomo é transferido para um translocon, elongação é retomada e proteína recém-sintetizada passa através da membrana para o lúmen do ER.
 liga-se ao peptídeo sinal e elongação é temporariamente paralisada. Etapa C: o ribossomo move-se para a membrana do ER, onde uma proteína de aportamento (docking) liga-se a SRP. Etapa D: ribossomo é transferido para um translocon, elongação é retomada e proteína recém-sintetizada passa através da membrana para o lúmen do ER.")
13
FIGURA 6.14Biossíntese de oligossacarídeos N-ligados na superfície do retículo endoplasmático.Etapa A: síntese começa sobre a face citoplasmática da membrana do ER com transferência de uma N-acetilglucosamina fosfato para um dolicol aceptor. Etapa B: formação da primeira ligação glicosídica açúcar-açúcar ocorre após transferência de um resíduo de N-acetilglucosamina. Etapa C: cinco resíduos de manose (de GDP-manose) são adicionados seqüencialmente, e o oligossacarídeo ligado a lipídeo é reorientado para a face luminal da membrana. Etapa D: resíduos adicionais de manose e (Etapa E) glicose são transferidos de intermediários ligados a dolicol. Dolicol-açúcares são gerados a partir de nucleosídeo difosfato-açúcares citoplasmáticos. Etapa F: o oligossacarídeo completo é transferido para um polipeptídeos nascente na superfície da membrana; peptídeo sinal pode já ter sido clivado nesse ponto
 são adicionados seqüencialmente, e o oligossacarídeo ligado a lipídeo é reorientado para a face luminal da membrana. Etapa D: resíduos adicionais de manose e (Etapa E) glicose são transferidos de intermediários ligados a dolicol. Dolicol-açúcares são gerados a partir de nucleosídeo difosfato-açúcares citoplasmáticos. Etapa F: o oligossacarídeo completo é transferido para um polipeptídeos nascente na superfície da membrana; peptídeo sinal pode já ter sido clivado nesse ponto.")
14
FIGURA 6. 15Estrutura de oligossacarídeos N-ligados
FIGURA 6.15Estrutura de oligossacarídeos N-ligados. Estruturas básicas de ambos os tipos de oligossacarídeos N-ligados são mostradas. Em ambos os casos, a estrutura deriva-se do oligossacarídeo inicial ligado ao dolicol por ação de glicosidases e glicosiltransferases. Note a variedade de ligações glicosídicas envolvidas nessas estruturas

15
FIGURA 6. 16Exemplos de estruturas de oligossacarídeos
FIGURA 6.16Exemplos de estruturas de oligossacarídeos. Estruturas 1-3 são oligossacarídeos N-ligados típicos dos tipos altamanose (1) e complexo (2, 3); note a estrutura do core comum do resíduo de asparagina da proteína até o primeiro ponto de ramificação. Estruturas 4-8 são oligossacarídeos O-ligados comuns, que podem ser bastante simples ou muito complexos. Note que embora a estrutura do core (GalNAc-Ser/Thr) seja diferente dos oligossacarídeos N-ligados, as extremidades podem ser muito semelhantes (p. ex., as estruturas 2 e 6, 3 e 7). Abreviaturas: Man = manose; Gal = galactose; Fuc = fucose; GlcNAc = N-acetilglucosamina; NANA = ácido N-acetilneuramínico (ácido siálico).Adaptado de Paulson, J. Trends Biochem. Sci. 14:272, 1989.
 e complexo (2, 3); note a estrutura do core comum do resíduo de asparagina da proteína até o primeiro ponto de ramificação. Estruturas 4-8 são oligossacarídeos O-ligados comuns, que podem ser bastante simples ou muito complexos. Note que embora a estrutura do core (GalNAc-Ser/Thr) seja diferente dos oligossacarídeos N-ligados, as extremidades podem ser muito semelhantes (p. ex., as estruturas 2 e 6, 3 e 7). Abreviaturas: Man = manose; Gal = galactose; Fuc = fucose; GlcNAc = N-acetilglucosamina; NANA = ácido N-acetilneuramínico (ácido siálico).Adaptado de Paulson, J. Trends Biochem. Sci. 14:272,")
16
FIGURA 6. 17Direcionamento de enzimas para lisossomos
FIGURA 6.17Direcionamento de enzimas para lisossomos. Glicoproteína N-ligada completa dobrada é liberada da membrana do ER e, antes do transporte para o aparelho de Golgi, glicosilases removem resíduos de glicose (Etapa 1). Um resíduo de manose pode também ser removido. Etapa 2: no aparelho de Golgi, uma glicosil transferase liga um ou, às vezes, dois resíduos de N-acetilglucosamina fosfato ao oligossacarídeo. Etapa 3: uma glicosidase remo-ve N-acetilglucosamina, deixando um ou dois resíduos de manose 6-fosfato no oligossacarídeo. A proteína é, então, reconhecida por um receptor de manose 6-fosfato e dire-cionada para vesículas que se destinam aos lisossomos.Adaptado de Kornfeld, R. e Kornfeld, S. Annu. Rev. Bio-chem. 54:631, 1985.
. Um resíduo de manose pode também ser removido. Etapa 2: no aparelho de Golgi, uma glicosil transferase liga um ou, às vezes, dois resíduos de N-acetilglucosamina fosfato ao oligossacarídeo. Etapa 3: uma glicosidase remo-ve N-acetilglucosamina, deixando um ou dois resíduos de manose 6-fosfato no oligossacarídeo. A proteína é, então, reconhecida por um receptor de manose 6-fosfato e dire-cionada para vesículas que se destinam aos lisossomos.Adaptado de Kornfeld, R. e Kornfeld, S. Annu. Rev. Bio-chem. 54:631,")
17
FIGURA 6.18Topologia de proteínas em membranas do retículo endoplasmático. Proteínas são mostradas em várias orientações com relação à membrana. Em (a) a proteína é ancorada à superfície luminal da membrana por um peptídeo sinal não-clivado. Em (b) a seqüência sinal não está na extremidade N-terminal; um domínio da proteína foi sintetizado antes da emergência do peptídeo sinal. Inserção da seqüência sinal interna, seguida por complementação da tradução, resultou em uma proteína com um domínio N-terminal citoplasmático, um segmento central que atravessa a membrana, e um domínio C-terminal no lúmen do ER. (c) Uma proteína com a orientação oposta: uma seqüência sinal N-terminal, que também poderia ter sido clivada pela peptidase sinal, resultou em extrusão de um segmento da proteína no lúmen do ER. Uma seqüência hidrofóbica de ancoragem permaneceu associada à membrana e impediu a continuação da passagem da proteína através da membrana, formando assim um domínio C-terminal citoplasmático. Em (d) vários sinais internos e seqüências de ancoragem permitem que vários segmentos da proteína fiquem orientados em ambos os lados da membrana.
 a proteína é ancorada à superfície luminal da membrana por um peptídeo sinal não-clivado. Em (b) a seqüência sinal não está na extremidade N-terminal; um domínio da proteína foi sintetizado antes da emergência do peptídeo sinal. Inserção da seqüência sinal interna, seguida por complementação da tradução, resultou em uma proteína com um domínio N-terminal citoplasmático, um segmento central que atravessa a membrana, e um domínio C-terminal no lúmen do ER. (c) Uma proteína com a orientação oposta: uma seqüência sinal N-terminal, que também poderia ter sido clivada pela peptidase sinal, resultou em extrusão de um segmento da proteína no lúmen do ER. Uma seqüência hidrofóbica de ancoragem permaneceu associada à membrana e impediu a continuação da passagem da proteína através da membrana, formando assim um domínio C-terminal citoplasmático. Em (d) vários sinais internos e seqüências de ancoragem permitem que vários segmentos da proteína fiquem orientados em ambos os lados da membrana..")
18
FIGURA 6. 19Maturação da pró-insulina humana
FIGURA 6.19Maturação da pró-insulina humana. Após clivagem nos dois pontos indicados por setas, resíduos de arginina 31, 32 e 65 e resíduo de lisina 64 são removidos para produzir insulina e peptídeo C.Redesenhado de Bell, G. I., Swain, W. F., Pictet, R., Cordell, B., Goodman, H. M. e Rutter, W. J. Nature 282:525, 1979

19
FIGURA 6.21Estrutura do colágeno, ilustrando (de cima para baixo) a regularidade da seqüência primária em uma hélice tipo II de poliprolina que gira para a esquerda; a tripla hélice que gira para a direita; a molécula de 300 nm; e a organização das moléculas em uma fibrila típica, dentro da qual moléculas de colágeno são unidas por ligações cruzadas FIGURA 6.22Modelo do proteassomo. Um segmento central 20S que inclui polipeptídeos diferentes é composto de quatro anéis heptaméricos empilhados, com um núcleo oco que inclui várias proteases com diferentes especificidades.Segmentos em forma de V em ambas as extremidades formam uma “tampa” ou cap para o cilindro e são responsáveis pelo reconhecimento de substrato ATP-dependente, desdobramento e translocação para o interior. A estrutura de cap superior está também em contato com o segmento central, mas é mostrada deslocada dele para mostrar o centro oco do cilindro.Adaptado de Rubin, D. e Finley, D. Curr. Biol. 5:854, Peters, J. -M. Trends Biochem. Sci. 19:377, 1994.

Apresentações semelhantes