Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Capítulo 9 Transcrição LGCM URLGA

2
9.1 Introdução O RNA é idêntico à fita de DNA codificadora LGCM URLGA

3
9.1 Introdução A transcrição começa quando a RNA polimerase se liga ao promotor O produto final da transcrição é chamado de transcrito primário LGCM URLGA

4
LGCM URLGA 9.1 Introdução Degradação – mRNA Clivagem – tRNA e rRNA
Procariotos Eucariotos Modificação das pontas – mRNA Clivagem – tRNA e rRNA LGCM URLGA

5
URLGA 9.1 Introdução Como a RNA polimerase encontra promotores no DNA?
Como proteínas regulatórias interagem com a RNA polimerase para ativar ou reprimir os passos da transcrição? URLGA

6
9.2 Como ocorre a trancrição
A transcrição ocorre em uma “bolha de transcrição”, onde o DNA é separado em duas fitas e a fita molde é usada na produção da fita de RNA O grupo 3’-OH reage com o trifosfato 5’. Há a perda de dois grupos fosfato ( e ) e o fosfato é usado na ligação da cadeia. URLGA
 e o fosfato é usado na ligação da cadeia. URLGA.")
7
9.2 Como ocorre a trancrição
A velocidade da reação é de nucleotídeos por segundo para RNA polimerase. A DNA polimerase tem uma taxa de 800 nucleotídeos por segundo A bolha de transcrição se move junto com a RNA polimerase URLGA

8
9.2 Como ocorre a trancrição
À medida que a RNA polimerase se move pela fita de DNA, ela desfaz a hélice na parte anterior e a refaz na parte posterior. O comprimento da bolha de transcrição é de 12 a 14 pb, mas a região híbrida DNA-RNA é menor Os últimos 25 nucleotídeos de RNA estão ligados a DNA ou à enzima. URLGA

9
9.3 A transcrição tem três estágios
Ligação da RNA pol a um promotor Elongação Parada e dissociação da RNA pol Transcrição Reconhecimento da fita começa com a ligação da RNA pol à fita dupla de DNA em um promotor para formar um complexo fechado. Então a fita de DNA é aberta formando um complexo aberto. URLGA

10
9.3 A transcrição tem três estágios
Durante a iniciação a enzima continua ligada ao promotor enquanto polimeriza as primeiras 9 bases. Durante a elongação a enzima se move pelo DNA estendendo a cadeia de RNA URLGA

11
9.3 A transcrição tem três estágios
A terminação envolve o reconhecimento do ponto ao qual não se deve mais adicionar bases à cadeia. Algumas mudanças no padrão da elongação podem acontecer quando a RNA pol pára. URLGA

12
9.4 RNA polimerase do fago T7
Cadeia polipeptídica simples < 100 kD Taxa de síntese: 200 nucleotídeos / segundo A RNA pol dos fagos T3 e T7 são pequenas e dão uma boa noção do aparato mínimo necessário. Ela é homóloga à DNA polimerase, onde o DNA se localiza em um palmo cercado por “dedos” e um “dedão” LGCM URLGA

13
9.4 RNA polimerase do fago T7
A RNA pol reconhece o DNA alvo ao se ligar a bases pela grande fissura da hélice. Durante a sua atividade o formato da enzima permanece essencialmente o mesmo. LGCM URLGA

14
9.5 Um modelo para o movimento da enzima é sugerido pela estrutura cristalizada
Estrutura comum entre RNA polimerase bacteriana e eucariótica: Canal na superfície: DNA – 16 a 25 bp. Enzima de levedura: 12 subunidades – estrutura de 10 Sítio catalítico entre duas subunidades maiores: adesão do DNA Subunidades 4 e 7: complexo que desagrega da enzima completa.

15
Um sítio catalítico contém um íon Mg2+.
O DNA é mantido no sítio catalítico pelas subunidades 1, 2 e 6. O DNA é forçado a se virar ao entrar no sítio devido a uma “parede” protéica. O comprimento da cadeia de RNA híbrida por outra proteína, ,chamada leme (rudder). Os nucleotídeos provavelmente entram na no sítio ativo por baixo, através de poros na estrutura.
. Os nucleotídeos provavelmente entram na no sítio ativo por baixo, através de poros na estrutura.")
16
A bolha de transcrição inclui 9 bp do híbrido DNA-RNA.
Onde o DNA se curva, as bases ali presentes são colocadas pra fora da hélice. Assim que a enzima se move, a base da fita molde se posiciona no sítio de entrada de nucleotídeos. O comprimento do híbrido é de 9bp e o lado 5’ do RNA deixa o DNA quando ele encontra a barreira protéica (rudder). Para a entrada do DNA no sítio ativo é necessária uma mudança conformacional na enzima: clamp
. Para a entrada do DNA no sítio ativo é necessária uma mudança conformacional na enzima: clamp.")
17
A estrutura da enzima sugere como a enzima mantém o contato com o substrato enquanto forma e quebra as ligações: Ponte – mudança conformacional durante translocação. Início: ponte estirada, adjacente ao sítio de entrada do nucleotídeo – interação com o novo nucleotídeo adicionado. Após o movimento da enzima, a ponte muda sua conformação, mantendo contato com o novo nucleotídeo e obstruindo o sítio de entrada. Final: retorno à conformação inicial

18
9.6 RNA polimerase bacteriana consiste de múltiplas subunidades
RNA polimerase melhor caracterizada. Responsável por quase toda a síntese de mRNA e de toda a síntese de rRNA e t RNA em eubactérias. A enzima completa (holoenzima) é formada por: subunidades b e b´: centro catalítico – b se liga à fita molde de DNA, ao RNA e aos ribonucleotídeos. subunidade a: necessária para reunião da enzima – reconhecimento de sequencias promotoras e interação com fatores de regulação. Subunidade s: reconhecimento de regiões promotoras.
 é formada por: subunidades b e b´: centro catalítico – b se liga à fita molde de DNA, ao RNA e aos ribonucleotídeos. subunidade a: necessária para reunião da enzima – reconhecimento de sequencias promotoras e interação com fatores de regulação. Subunidade s: reconhecimento de regiões promotoras.")
19
O canal do DNA se localiza entre as subunidades b e b´
O canal do DNA se localiza entre as subunidades b e b´. As fitas são desenroladas no sítio ativo, onde é feita a síntese de RNA. As subunidades b e b´ interagem com com DNA em diversos sítios diversos do sítio ativo e com RNA – estabilização das fitas desnaturadas. Rimfapicina: bloqueio de RNA polimerase – ligação ao sítio de elongação do RNA na subunidade b. RNA polimerase: além de catalisar síntese, supervisiona pareamento e faz ligações fosfodiester entre o DNA e o RNA. Todas as subunidades ativas durante o elongamento da cadeia são necessárias para início e término: diferenças na dependência de peptídeos adicionais durante as fases. Complexidade enzimática reflete em parte a necessidade de interação com fatores regulatórios.

20
9.7 RNA polimerase consiste do núcleo da enzima mais o fator s
A holoenzima pode ser dividida em dois componentes: o núcleo e o fator s. Apenas a holoenzima pode iniciar a transcrição O fator s garante que a enzima se ligou estavelmente ao DNA em uma região promotora. O fator se desliga da enzima após as primeira 8-9 bases. O núcleo pode sintetizar o RNA à partir do molde, mas não iniciar a transcrição em um sítio correto. O fator s produz a modificação na afinidade na RNA polimerase por DNA que é alta. No entanto, o fator s confere habilidade de reconhecer sítios de ligação específicos, promovendo a ligação da holoenzima aos promotores firmemente.

21
9.8 A associação com o fator s muda na iniciação
Formação de um complexo fechado (DNA em fita dupla) – KB Formação do complexo aberto (desnaturação de parte da fita dupla ligada a enzima). Reação rápida – s. Incorporação dos primeiros 2 nucleotídeos e formação da ligação fosfodiester – complexo ternário. Ocorrem ciclos de iniciação abortiva. Sucesso da iniciação: liberação de s e movimento da enzima núcleo pela fita molde.
 – KB. Formação do complexo aberto (desnaturação de parte da fita dupla ligada a enzima). Reação rápida – s. Incorporação dos primeiros 2 nucleotídeos e formação da ligação fosfodiester – complexo ternário. Ocorrem ciclos de iniciação abortiva. Sucesso da iniciação: liberação de s e movimento da enzima núcleo pela fita molde.")
22
Ao se ligar ao DNA, a conformação da RNA polimerase sofre mudanças
Ao se ligar ao DNA, a conformação da RNA polimerase sofre mudanças. Essas mudanças identificam 2 formas do complexo: Iniciação: enzima cobre de bp; Transição da iniciação para elongação: 60bp – sequencia promotora inicial envolvida no reconhecimento inicial pela RNA polimerase. Após adição de 15 a 20 bases, uma nova mudança ocorre na enzima – elongação. Nem todos os complexos da RNA polimerase liberam o fator s durante a elongação (70%).
.")
23
Tendem a se concentrar em posições mais conservadas
9.13 – A eficiência do promotor pode ser aumentada ou diminuída por mutação Mutações em regiões promotoras Alteração no nível de expressão dos genes que elas controlam expressão “down” mutações Tendem a se concentrar em posições mais conservadas Freqüentemente aumentam a homologia com uma das seqüências consenso ou tornam a distância entre elas mais próximas a 17 pb. expressão “up” mutações

24
Determinação do efeito absoluto das mutações promotoras
Medida da afinidade da RNA polimerase aos promotores selvagem e mutante Investigar o estágio no qual a mutação influencia a capacidade do promotor Alteração da afinidade para ligação a RNA polimerase Alteração na etapa de iniciação Alteração da influência de um fator auxiliar

25
Domínio de reconhecimento
Mutações “down” na seqüência em –35 reduzem a taxa de formação do complexo fechado, mas não inibem a conversão para o complexo aberto; Mutações “down” na sequência em –10 não afetam a formação inicial do complexo fechado, mas tornam mais lenta sua conversão para a forma aberta. Domínio de reconhecimento Seqüências de A e T Domínio de desenrolamento

26
Seqüências imediatamente ao redor do ponto de início influenciam o evento de iniciação da transcrição; Região inicialmente transcrita (+1 a +30) influenciam a taxa na qual a RNA polimerase desliga-se do promotor, tendo efeito na força do promotor.
 influenciam a taxa na qual a RNA polimerase desliga-se do promotor, tendo efeito na força do promotor.")
27
9.14 – A RNA polimerase liga-se a uma face do DNA
Caracterização da capacidade da RNA polimerase de reconhecer DNA Footprinting

28
Identificação dos pontos nos quais a RNA polimerase realmente entra em contato com o promotor
Modificação da técnica: - Modificação de bases do DNA antes de sua ligação à RNA polimerase. Identificação dos sítios que a enzima deve reconhecer para se ligar ao DNA - Modificação do complexo RNA pol – DNA proteção contra a modificação Identificação de sítios que fazem contato com a RNA pol (sítios de reconhecimento mais algumas posições adicionais)
")
29
Maioria dos pontos de contato
Posições comuns para diferentes promotores Mecanismo comum para a ligação, independente da presença de uma base particular

30
9.15 – Superanelamento é uma característica importante da transcrição
A RNA pol pode iniciar a transcrição mais eficientemente in vitro quando a fita molde está superanelada. Presumivelmente devido ao fato dessa estrutura requerer menos energia livre para a separação inicial das fitas de DNA no complexo de iniciação. Alguns promotores são influenciados pelo grau de superanelamento Mais fáceis de separar Mais difíceis de separar Determinado pela seqüência Ou pela localização

31
Afeta a estrutura local do DNA
Para cada volta da hélice atravessada pela RNA pol, +1 volta é gerada a frente e –1 atrás. Afeta a estrutura local do DNA Girase Topoisomerase

32
9.16 – Substituição de fatores sigma pode controlar iniciação
Existe divisão de trabalho entre o cerne da enzima, que faz a elongação durante a transcrição e um fator sigma, envolvido da seleção do sítio onde ocorrerá ligação da holoenzima. Haverá mais de um tipo de fator sigma, cada um específico para uma classe diferente de promotores?

33
E. Coli usa fatores sigma alternativos para responder a mudanças ambientais.

34
Variação na temperatura é um tipo comum de obstáculo para os seres vivos.
Muitos organismos, eucarióticos e procarióticos, respondem de uma forma semelhante. Com o aumento da temperatura, a síntese de proteínas normalmente produzidas é cessada ou diminuída, e um novo conjunto de proteínas é sintetizado. Estas atuam protegendo a célula contra estresses ambientais, sendo grande parte destas chaperones.

35
- A resposta ao choque térmico é conferida por um aumento do fator sigma 32, quando a temperatura aumenta, e um decréscimo deste fator quando a temperatura volta a abaixar. O sina básico que induz a produção de sigma 32 é o acúmulo de proteínas não moldadas (parcialmente desnaturadas). O sigma 32 é instável, o que permite que sua quantidade na célula aumente e diminua rapidamente. Sigma 70 e sigma 32 podem competir pelo cerne da enzima disponível, assim o conjunto de genes expressos durante o choque térmico depende de um balanço entre esses fatores.
. O sigma 32 é instável, o que permite que sua quantidade na célula aumente e diminua rapidamente. Sigma 70 e sigma 32 podem competir pelo cerne da enzima disponível, assim o conjunto de genes expressos durante o choque térmico depende de um balanço entre esses fatores.")
36
- Outro grupo de gene regulados por temperatura é controlado pelo fator sigma E, que responde a aumentos mais extremos. O acúmulo de proteínas no periplasma ativa a protease DegS. A clivagem da porção C-terminal de RseA por DegS ativa a protease YaeL. Esta cliva a região N-terminal de RseA, liberando o fator sigma E

37
No caso de escassez de nitrogênio, E
No caso de escassez de nitrogênio, E. coli usa o fator sigma 54, o qual normalmente está em baixas quantidades e é ativado quando não há amônia no meio. O objetivo é transcrever genes que permitam a utilização de fontes alternativas de nitrogênio. Análogos desse gene foram encontrados em várias outras bactérias. Esse mecanismo foi conservado durante a evolução Outro análogo de genes de E. coli (fator sigma F) é o fator sigma D de B. subtilis, que controla genes ligados à mobilidade e ao flagelo.
 é o fator sigma D de B. subtilis, que controla genes ligados à mobilidade e ao flagelo.")
38
Um aspecto importante para os promotores de cada enzima é que eles possuem o mesmo tamanho e a mesma localização relativa ao códon de iniciação. Apenas as regiões próximas ao -35 e -10 possuem conservação (exceto o sigma 54, -24 e -12) - Uma enzima contendo determinado fator sigma só ligará a promotores com a seqüência -35 e -10 que ela reconhece
 - Uma enzima contendo determinado fator sigma só ligará a promotores com a seqüência -35 e -10 que ela reconhece.")
39
9.17 – Fatores sigma entram diretamente em contato com o DNA
Os fatores sigmas se ligam ao promotor nas regiões -35 e -10. Este fato pode ser constatado por mutações no fator sigma que suprimem mutações nas seqüências consenso. Comparações entre vários sigmas de bactérias identificaram regiões que foram conservadas

40
O uso de motivos alfa-hélice em proteínas para reconhecer seqüências de DNA é comum.
Aminoácidos separados por 3 a 4 resíduos se situam na mesma face de uma alfa-hélice, interagindo com pares de bases adjacentes.

41
A região N-terminal do sigma 70 tem um importante papel regulatório.
Ao ser removida, a proteína então mais curta se liga mais especificamente a promotores Isso sugere que essa região se comporta como um domínio de auto-inibição. A região obstrui o domínio de ligação a DNA quando o fator está livre. Esse domínio só é liberado quando o fator está ligado à holoenzima.

42
9.18 – Fatores sigma podem se organizar em cascatas
Mudanças de um conjunto de genes expressos para outro é um aspecto comum de infecção por bacteriófago. Quando o fago infecta, ocorre uma mudança no padrão de transcrição durante seu ciclo infectivo. Um exemplo conhecido é o de infecção de B. subtilis pelo fago SPO1. O ciclo de infecção do SPO1 passa por 3 estágios.

43
O padrão de transcrição cria uma cascata, na qual a enzima do hospedeiro transcreve um gene que será necessário para a transcrição de outro gene posterior Isso faz com que a enzima não transcreva mais os genes do hospedeiro, mas sim do fago.

44
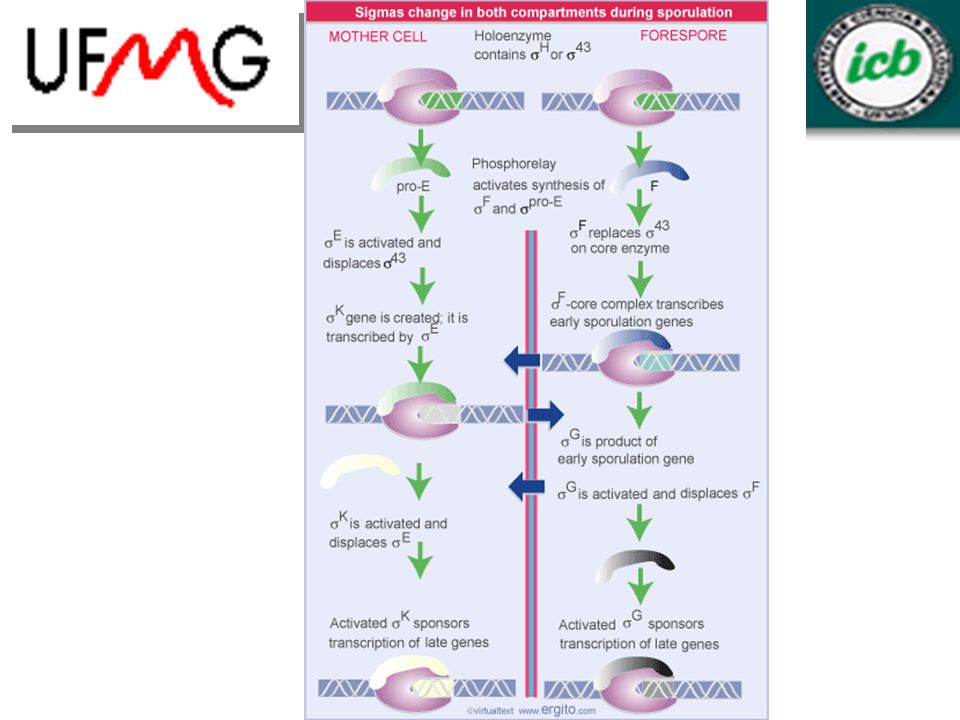
9.19 Esporulação é controlada por fatores sigma
crescimento logarítmico para devido a falta de nutrientes, acionando esporulação septo prende parte do cromossomo que vai ser deslocado por translocase (SpoIIIE) processo dura aprox. 8 hrs. Envolve mudanças drásticas nas atividades biossintéticas da bactéria
 processo dura aprox. 8 hrs. Envolve mudanças drásticas nas atividades biossintéticas da bactéria.")
45
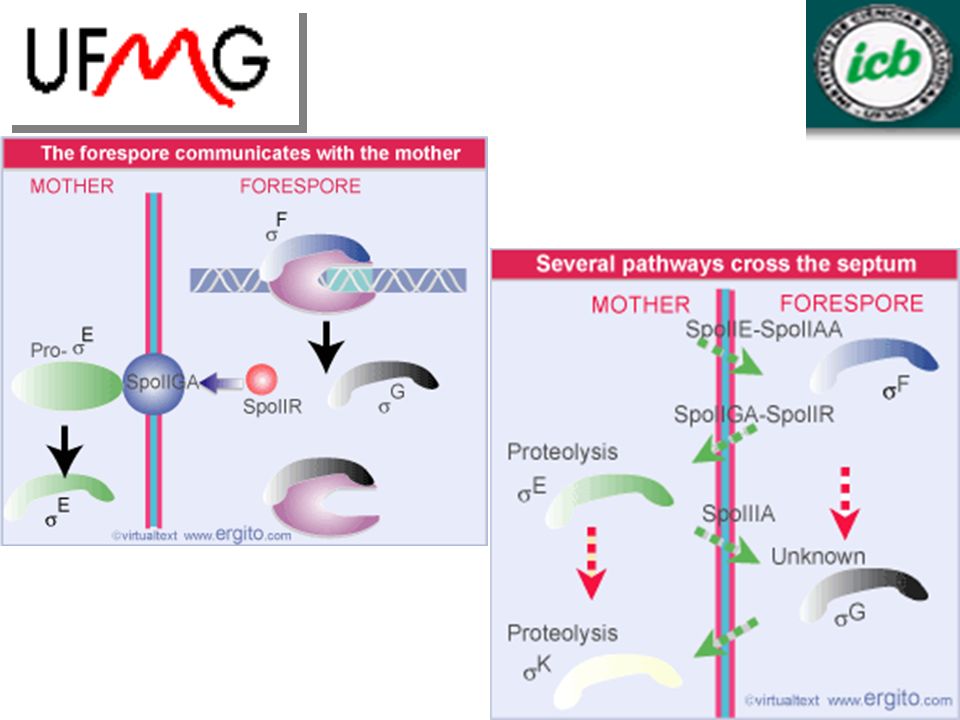
Novas formas da RNA polimerase são ativadas em células no processo de esporulação
fator sigma existente é substituído por novo fator que transcreve conjunto diferente de genes comunicação entre os compartimentos coordena mudanças na célula F é inibido por fator anti-sigma (SpoIIAB) SpoIIE desfosforila SpoIIAA que libera F ativação de F é o começo da esporulação F é produzido em pequena quantidade
 SpoIIE desfosforila SpoIIAA que libera F. ativação de F é o começo da esporulação. F é produzido em pequena quantidade.")
48
9.20 RNA polimerase bacteriana termina em sítios distintos
terminação requer quebra de todas as pontes de hidrogênio RNA-DNA para dúplex de DNA seja reformado seqüências que já foram transcritas pela RNA polimerase são responsáveis pela terminação muitos terminadores requerem formação de grampo na estrutura secundária do RNA transcrito

49
paralelo entre iniciação e terminação:
A eficiência dos terminadores varia muito: evento de terminação pode ser evitado por fator ancilar específico eventos de terminação podem ser usados como controle da expressão gênica paralelo entre iniciação e terminação: interação entre dúplex de DNA e RNA polimerase reconhecimento de sinais no transcrito e seqüências no DNA

50
9.21 Existem dois tipos de terminadores em E. coli
Terminador rho-dependente Terminador denominado intrínseco quando RNA polimerase não precisa de nenhum fator adicional para a terminação in vitro ~1100 seqüências em E. coli apresentam características de terminadores intrínsecos mutações pontuais na região da haste impedem terminação

51
Grampo pode diminuir ou parar síntese de RNA dando oportunidade para a terminação
região rica em U desestabiliza híbrido RNA-DNA seqüência do grampo e extensão da região rica em U influenciam na eficiência da terminação parâmetros adicionais influenciam na interação com a RNA polimerase: seqüências upstream e downstream

52
9.22 Como funciona o fator rho?
Proteína essencial em E. coli: funciona somente no estágio de terminação Hexâmero de ~275kD com subunidades idênticas: domínio de ligação ao RNA e domínio de hidrólise do ATP Helicase ATP-dependente: passagem de nucleotídeos pelo buraco formado por hexâmero terminadores rho-dependentes representam metade dos terminadores de E. coli

53
Função chave é a atividade helicase, energia fornecida por hidrólise de ATP RNA-dependente
Atividade da ATPase fornece energia para mover-se ao longo do RNA Pausa é importante pois fornece tempo para eventos necessários terminação por fator rho pode ser impedida por ribossomo em tradução

54
Em alguns casos, mutação nonsense em unidade transcricional impede a transcrição dos genes seguintes: POLARIDADE

55
Mutações em rho tem influência variada na terminação
Terminadores diferentes requerem níveis diferentes do fator rho para terminação Algumas mutações em rho podem ser suprimidas por mutações em outros genes método para identificar proteínas que interagem com rho

Apresentações semelhantes