Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Origens e síntese do acetil-CoA, catabolismo dos aminoácidos cetogénicos e relação com o ciclo de Krebs

2
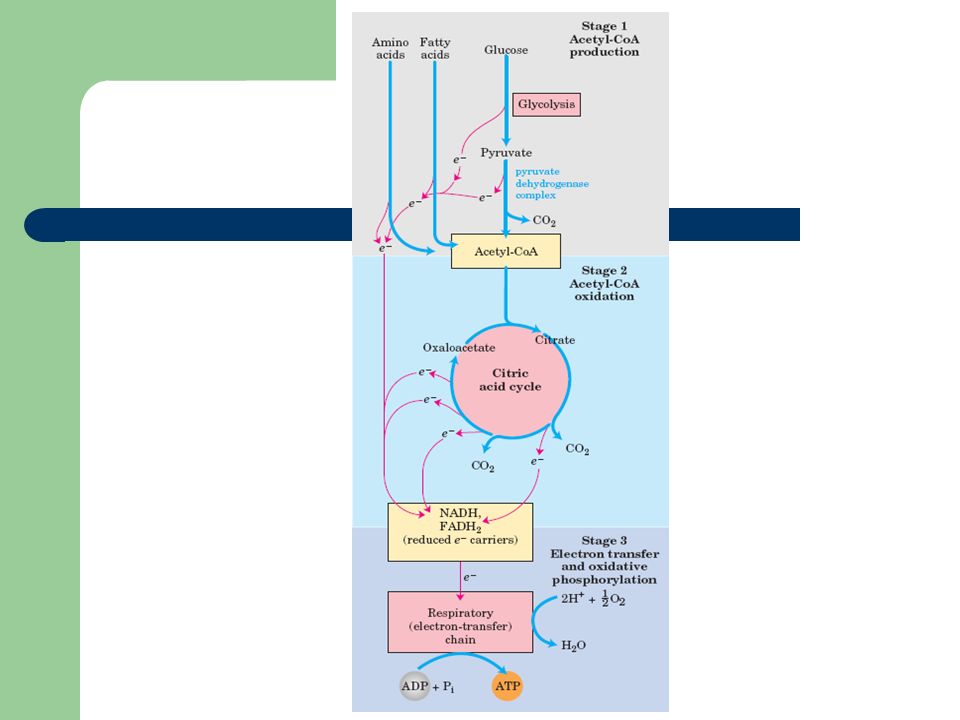
Origens e síntese do Acetil-CoA Acetil CoA GlicogénioTriacilgliceróisProteína Glicose Piruvato Ácidos gordos livres Aminoácidos Proteólise Desaminação oxidativa Glicogenólise Glicólise Descarboxilação Oxidativa Lipólise β -oxidação

3
Acetil CoA GlicogénioTriacilgliceró is Proteína Glicose Piruvato Ácidos gordos livres Aminoácidos Proteólise Desaminação oxidativa Glicogenólise Glicólise Descarboxilaç ão Oxidativa Lipólise β -oxidação

4
A acetil-CoA é formada a partir da descarboxilação oxidativa do piruvato, realizada sequencialmente pela piruvato desidrogenase –PDH (complexo multienzimático de 3 enzimas), na matriz mitocondrial: Desidrogenase pirúvica (grupo prostético TPP) Dihidrolipoiltranscetilase (grupo prostético Lipoamida) Dihidrolipoildesidrogenase (grupo prostético FAD) – …e 5 coenzimas: Tiamina pirofosfato (TPP) – reage com o piruvato Lipoamida – aceita grupo acetil e transfere-o para o CoA CoA – aceita grupo acetil FAD – aceita equivalentes redutores NAD+ – aceita equivalentes redutores Síntese de Acetil-CoA: 1-Descarboxilação oxidativa do piruvato
, na matriz mitocondrial: Desidrogenase pirúvica (grupo prostético TPP) Dihidrolipoiltranscetilase (grupo prostético Lipoamida) Dihidrolipoildesidrogenase (grupo prostético FAD) – …e 5 coenzimas: Tiamina pirofosfato (TPP) – reage com o piruvato Lipoamida – aceita grupo acetil e transfere-o para o CoA CoA – aceita grupo acetil FAD – aceita equivalentes redutores NAD+ – aceita equivalentes redutores Síntese de Acetil-CoA: 1-Descarboxilação oxidativa do piruvato")
5
Reacção global: Reacção altamente exergónica É irreversível Ácidos gordos não podem gerar directamente glícidos

6
2 - β-oxidação

7
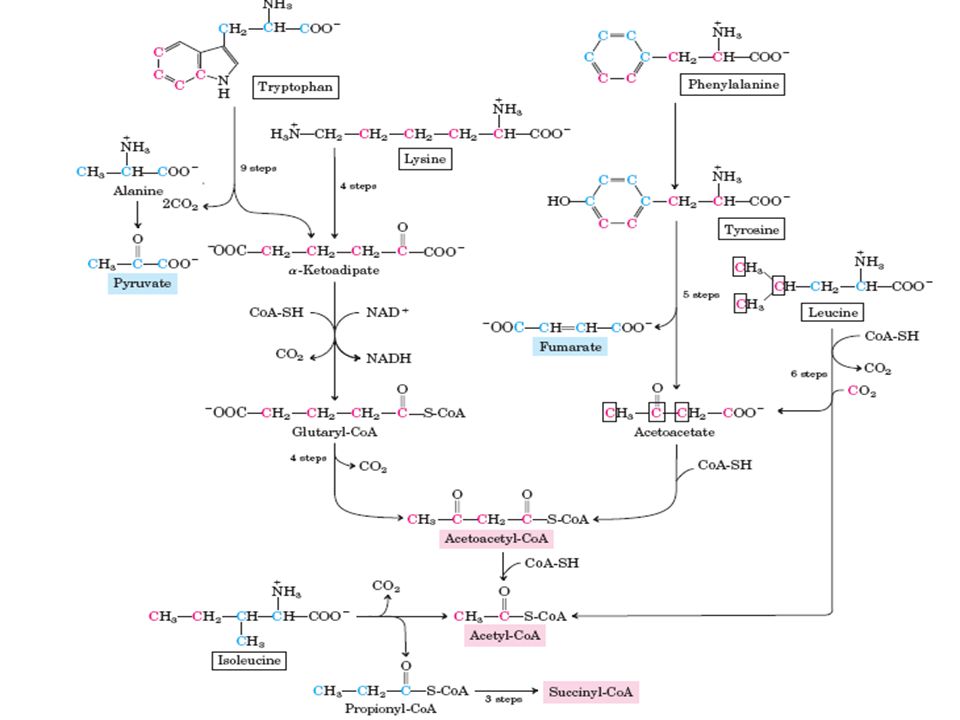
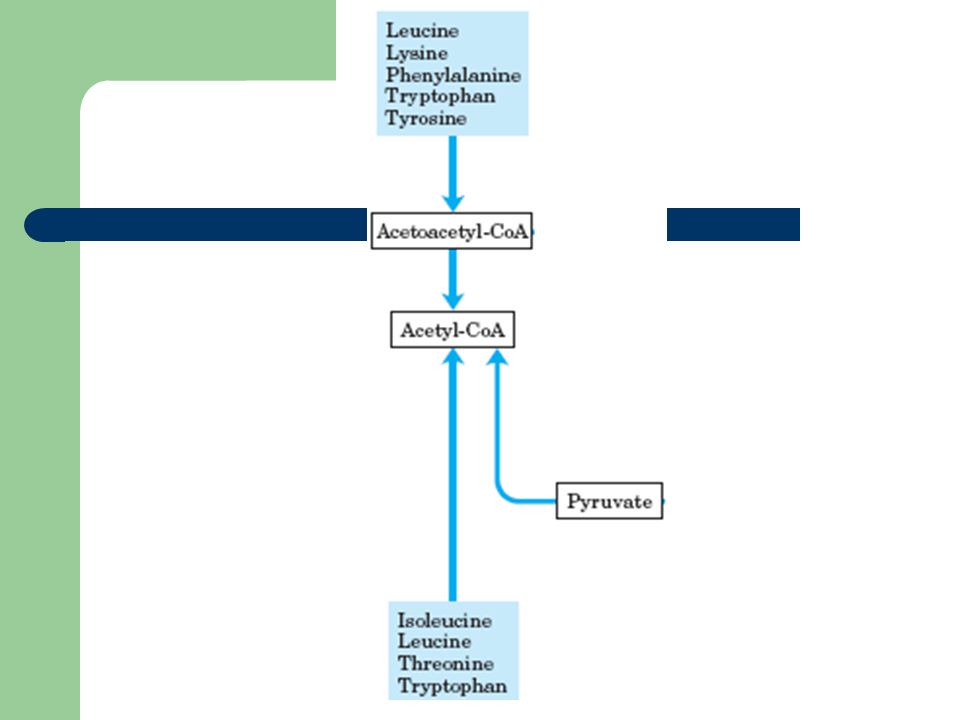
3 – Desaminação oxidativa no decurso do seu catabolismo desdobram-se, levando à formação de Acetil-CoA e Acetoacetil-CoA Aminoácidos Cetogénicos

8
......

11
Acetoacetil-CoA (1) Tirosina transaminação (Glutamato) Para-hidroxifenilpiruvato Fumaril-acetoacetato hidrolisado (Fumarato) Acetoacetato Acetoacetil-CoA
 Tirosina transaminação (Glutamato) Para-hidroxifenilpiruvato Fumaril-acetoacetato hidrolisado (Fumarato) Acetoacetato Acetoacetil-CoA")
12
Acetoacetil-CoA (2) Lisina Leucina Fenilalanina (que se converte em tirosina): fenilalanina + tetrahidrobiopterina + O2 → TIROSINA + dihidrobiopterina + H2O ( por acção da hidroxílase da fenilalanina) Triptofano Glutatil-CoAAcetoacetil-CoA
 Lisina Leucina Fenilalanina (que se converte em tirosina): fenilalanina + tetrahidrobiopterina + O2 → TIROSINA + dihidrobiopterina + H2O ( por acção da hidroxílase da fenilalanina) Triptofano Glutatil-CoAAcetoacetil-CoA")
13
Acetil-CoA Conversão de Acetoacetil-CoA em Acetil-CoA

14
Acetil-CoA (1) Isoleucina Perda dos grupos α-amina em reacções de transaminação, formando α-cetoácidos ramificados. α-cetoácidos ramificado + CoA + NAD+ → acetil-CoA ramificado + CO2 + NADH + Propionil-CoA ( cisão tiolítica); Leucina Perda dos grupos α-amina em reacções de transaminação, formando α-cetoácidos ramificados α-cetoácidos ramificado + CoA + NAD+ → acil-CoA ramificado + CO2 + NADH Acil-CoA Acetoacetato + Acetil-CoA ( por cisão)
; Leucina Perda dos grupos α-amina em reacções de transaminação, formando α-cetoácidos ramificados α-cetoácidos ramificado + CoA + NAD+ → acil-CoA ramificado + CO2 + NADH Acil-CoA Acetoacetato + Acetil-CoA ( por cisão).")
15
Acetil-CoA (2) Treonina Treonina desidrogenase 2-amino-3-cetobutirato Acetil-CoA ligase
 Treonina Treonina desidrogenase 2-amino-3-cetobutirato Acetil-CoA ligase")
16
Triptofano (gera-se através da hidrólise de um intermediário do catabolismo da 3- hidrocinurenina) Libertação de alanina e 3-hidroxiantranilato Acetil-CoA Acetil-CoA (3)
 Libertação de alanina e 3-hidroxiantranilato Acetil-CoA Acetil-CoA (3)")
17
Ciclo de Krebs Etapas e intervenientes

18
Ciclo de Krebs (ciclo dos ácidos tricarboxílicos, ciclo do ácido cítrico) Hans Krebs (1900-1981) Realiza-se na matriz mitocondrial, porque as principais fontes de acetil-CoA estão na mitocôndria Ciclo anfibólico Objectivo Oxidação completa da acetil-CoA em CO 2 ; Formação de equivalentes redutores. Gerar ATP na cadeia de transporte de electrões e fosforilação oxidativa.

20
Substracto: acetil-CoA Produtos: 2 CO 2, 1 GTP (ATP), 3 NADH, 1 FADH 2
, 3 NADH, 1 FADH 2")
22
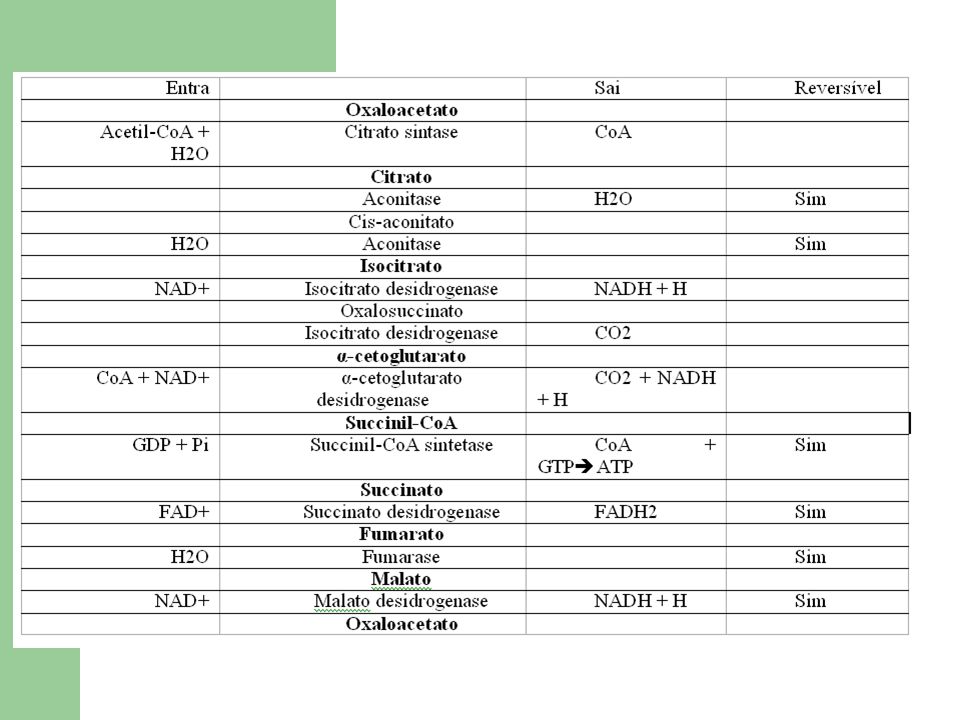
1ª reacção: formação de citrato Citrato sintase: cadeia polipéptidica com 2 domínios diferentes, com o centro activo entre estes. Formação de um composto intermédio bastante energético ( citroil-CoA), que rapidamente se transforma em citrato, libertando CoA. Reacção de condensação altamente exergónica. Formas de regulação da enzima dependente da quantidade dos substractos.
, que rapidamente se transforma em citrato, libertando CoA. Reacção de condensação altamente exergónica. Formas de regulação da enzima dependente da quantidade dos substractos..")
23
2ª reacção: formação de isocitrato Reacção catalisada pela enzima aconitase ( ou aconitase hidratase). Composto por 2 reacções envolvendo a perca e recuperação de H 2 O. Formação de cis-aconitato é endergónica. Logo cis-aconitato citrato exergónica, com tendência para se dar. No entanto o isocitrato está em baixa concentração na célula e consome-se rapidamente na próxima reacção, conseguindo a aconitase captar H 2 O,sendo necessário Fe2+ como co-factor.

24
3ª reacção: formação de α-cetoglutarato Este conjunto de 3 reacções (descarboxilação oxidativa) é primeiramente catalizado pela enzima isocitrato desidrogenase ( requer a presença de NAD+). Essencial a ligação de Mn2+ ou Mg2+ ao grupo carbonilo do composto intermédio oxalosuccinato para que se forme o produto final. Nesta reacção é produzido NADH e CO 2.

25
4ª reacção: formação de succinil-CoA Descarboxilação oxidativa catalizada pelo complexo multienzimático α-cetoglutarato desidrogenase (α-cetoglutarato, dihidrolipoil transsuccinilase e dihidrolipoil desidrogenase), na presença de coenzimas e grupos proestéticos (tiamina pirofosfato, ácido lipoico, CoA, FAD e NAD+), activada pelo Ca2+. Forma-se, igualmente NADH e CO 2. A ligação S-CoA conserva a energia. Na presença de glutamato desidrogenase, um intermediário de α- cetoglutarato pode ser redutoramente aminado e deixar o ciclo ( na presença de NADH e amónia) dando origem ao glutamato.
 dando origem ao glutamato..")
26
5ª reacção: formação de succinato Na presença do nucleósido difosfato cinase. Permitindo a fosforilação a nível do substracto de GDP a GTP. A succinil-CoA sintetase (constituida por 2 subunidades) convertendo o substracto em succinato, quebrando uma ligação bastante energética. Conservando o carácter de alta energia da ligação tiol-éster. Equação reversível comATP e GTP são energeticamente equivalentes
 convertendo o substracto em succinato, quebrando uma ligação bastante energética. Conservando o carácter de alta energia da ligação tiol-éster. Equação reversível comATP e GTP são energeticamente equivalentes.")
27
6ª reacção: formação de fumarato Succinato oxidado a fumarato pela succinato desidrogenase. FAD ligado covalentemente Intimamente ligada na membrana mitocondrial interna. Complexo II da cadeia respiratória ( passagem de electrões) Malonato ( semelhante ao succinato) é um inibidor desta reacção podendo ligar-se ao succinato desidrogenase. Bloquear o Ciclo de Krebs
 Malonato ( semelhante ao succinato) é um inibidor desta reacção podendo ligar-se ao succinato desidrogenase. Bloquear o Ciclo de Krebs.")
28
7ª reacção: formação de malato O fumarato é hidratado a malato ( mais concretamente L-malato) pela enzima fumarase. Só no fumarato ( devido a ligação dupla) é que esta reacção ocorre.
 é que esta reacção ocorre..")
29
8ª reacção: formação de oxaloacetato Catalizada pela enzima malato desidrogenase na presença de NAD+, havendo formação de NADH. Reacção endergónica no sentido directo Baixa concentração de oxaloacetato na célula (novo ciclo) e a oxidação de NADH a NAD+ pela respiração mitocondrial obrigam a reacção a progredir neste sentido. Oxaloacetato Aspartato ( transminação) Piruvato ( descarboxilação)
 e a oxidação de NADH a NAD+ pela respiração mitocondrial obrigam a reacção a progredir neste sentido. Oxaloacetato Aspartato ( transminação) Piruvato ( descarboxilação).")
30
Relembrar… A completa oxidação da acetil-CoA em CO2 e H2O conserva energia O ciclo serve como via oxidativa terminal para a maioria dos combustíveis metabólicos

31
Ciclo de Krebs Características anfibólicas, energéticas e regulação

32
Características Energéticas Equação geral do ciclo

33
Características Energéticas Rendimento energético: 3 NADH 1 FADH2 1 GTP/ATP

34
Características Energéticas Moléculas de NADH e FADH2, reduzidas nas etapas oxidativas Gerando mais ATP durante a fosforilação oxidativa Transporte de electrões para a cadeia respiratória

35
Características anfibólicas O ciclo de Krebs é um ciclo anfibólico Participa em reacções de catabolismo e anabolismo

36
Características anfibólicas Catabolismo O ciclo de Krebs participa do catabolismo oxidativo de: Ácidos gordos Oses Aminoácidos que servem como substrato em diferentes etapas do ciclo.

37
Características anfibólicas Anabolismo Vários intermediários do ciclo servem como precursores em reacções de biossíntese.

38
Características anfibólicas

39
Anabolismo As concentrações dos intermediários têm de se manter constantes, caso contrário o ciclo não decorre normalmente. Como tal, os intermediários têm de ser repostos por outros compostos, através de reacções anapleróticas.reacções anapleróticas

40
Regulação A regulação faz-se a dois níveis: antes do início do ciclo, na conversão do piruvato em Acetil-CoA; durante o ciclo, nas 3 etapas exergónicas.

41
Regulação Regulação da conversão de piruvato em Acetil-CoA Pode ser: Alostérica Covalente Positiva - presença de AMP, CoA, Ca2+, NAD+ Negativa – presença de ATP, ácidos gordos, NADH e Acetil-CoA Acção na presença de ATP sobre uma das subunidades da enzima E1 do complexo PDH

42
Regulação Regulação do ciclo Ocorre principalmente em 3 etapas consideradas limitantes: 1- Síntese do citrato; 2- Conversão do isocitrato em α-cetoglutarato; 3- Conversão do α-cetoglutarato em succinil-CoA. Pode também ocorrer nas outras etapas, devido à concentração de substratos e de produtos finais.

Apresentações semelhantes







 ou ANAeróbia (outras substâncias);>")
>")










