Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Comunidades: teoria da diversidade (I) Curso Especialização em Gestão de Recursos Hídricos
Pinto-Coelho, R.M. Departamento de Biologia Geral Instituto de Ciências Biológicas – ICB UFMG

2
Introdução O estudo da biodiversidade é um capítulo básico na Ecologia de comunidades. Refere-se ao estudo das relações quantitativas entre riqueza e abundância de espécies dentro das comunidades. Wallace (1878) foi um dos primeiros a reconhecer a importância da diversidade na caracterização das comunidades. Ele verificou que, nos trópicos, a diversidade das comunidades é muito alta. MacArthur & Wilson verificaram que ilhas longínquas e ou pequenas têm menos espécies do que ilhas próximas e extensas. Como podemos explicar estas tendências? Como estão distribuídas as abundâncias de diferentes espécies em uma comunidade? As informações básicas usadas são as seguintes: S: número total de espécies. N: número total de indivíduos. N1, N2, N3,..., Ni: números de indivíduos da cada espécie
 foi um dos primeiros a reconhecer a importância da diversidade na caracterização das comunidades. Ele verificou que, nos trópicos, a diversidade das comunidades é muito alta. MacArthur & Wilson verificaram que ilhas longínquas e ou pequenas têm menos espécies do que ilhas próximas e extensas. Como podemos explicar estas tendências Como estão distribuídas as abundâncias de diferentes espécies em uma comunidade As informações básicas usadas são as seguintes: S: número total de espécies. N: número total de indivíduos. N1, N2, N3,..., Ni: números de indivíduos da cada espécie.")
3
Diversidade é mais do que riqueza de espécies...
Na ilustração ao lado (extraída de Magurran, 1996, pág. 8), quando comparados as áreas A e B, A área B seria mais diversa porque possui três espécies de mariposas enquanto A tem apenas uma. Numa segunda situação, não existe qualquer diferença entre o número total de espécies entre as áreas C e D. O sítio C possui quatro espécies sendo que cada uma delas aparece com três indivíduos. A área D também possui quatro espécies sendo que uma delas é particularmente abundante, tendo nove indivíduos e as demais com apenas uma ocorrência. Dessa forma, embora as áreas C e D tenham a mesma riqueza em espécies de mariposas, a maior equitatividade observada em C faz dessa área uma área mais diversa. Embora o exemplo seja simples, ele ilustra o tipo de quantificação que será feita nos modelos a serem discutidos no capítulo.
, quando comparados as áreas A e B, A área B seria mais diversa porque possui três espécies de mariposas enquanto A tem apenas uma. Numa segunda situação, não existe qualquer diferença entre o número total de espécies entre as áreas C e D. O sítio C possui quatro espécies sendo que cada uma delas aparece com três indivíduos. A área D também possui quatro espécies sendo que uma delas é particularmente abundante, tendo nove indivíduos e as demais com apenas uma ocorrência. Dessa forma, embora as áreas C e D tenham a mesma riqueza em espécies de mariposas, a maior equitatividade observada em C faz dessa área uma área mais diversa. Embora o exemplo seja simples, ele ilustra o tipo de quantificação que será feita nos modelos a serem discutidos no capítulo.")
4
Riqueza e abundância relativa de espécies em uma comunidade
A medida mais simples é o número de espécies encontradas numa comunidade. Devem ser excluídas as espécies acidentais, exóticas e eventuais imigrantes ocasionais. Embora seja a maneira mais usual de se iniciar estudo da estrutura de uma comunidade, tal critério tem a limitação básica de não levar em consideração os padrões de abundância das espécies. Tabela: Abundâncias e riqueza em espécies em duas comunidades hipotéticas. As uas comunidades tem a mesma riqueza mas as abundâncais são muito diferentes entre si. A comunidade II é intuitivamente mais diversa. O conceito central dessa diferença é a noção de abundância relativa, pi, que é uma razão entre a abundância de cada espécie (ni) e o número total da amostra (N). Lembrar, no entanto, que tanto pi quanto N variam enormemente tanto com o esforço amostral quanto com a metodologia de coletas. No exemplo acima para a comunidade I temos na = 0,99 e nb= 0,01 enquanto na comunidade II tais números são 0,50 e 0,50, respectivamente.
 e o número total da amostra (N). Lembrar, no entanto, que tanto pi quanto N variam enormemente tanto com o esforço amostral quanto com a metodologia de coletas. No exemplo acima para a comunidade I temos na = 0,99 e nb= 0,01 enquanto na comunidade II tais números são 0,50 e 0,50, respectivamente.")
5
Diversidade tem escala ! - Diversidades local e regional
Suponha que os habitats encontrados nas regiões A e B possam suportar no máximo 10 espécies (tabela abaixo). Na região A as 10 espécies são generalistas e podem habitar todos os cinco habitats disponíveis. Na região B, as espécies são especialistas e só podem habitar num único habitat. Por conseguinte, toda a região poderá conter 50 espécies já que em cada habitat uma comunidade específica será encontrada. O exemplo ilustra os dois tipos de diversidade: a diversidade alfa ou local e a diversidade beta ou regional. As regiões A e B têm a mesma diversidade alfa, mas a região B tem uma diversidade beta maior. A diversidade alfa depende da capacidade do habitat em suportar um certo número de espécies o que é função do grau de competição entre as populações das diferentes espécies. A diversidade regional depende do grau de substituição de umas espécies pelas outras em diferentes habitats. Tabela 1: Dados hipotéticos de riqueza em espécies em duas regiões com diferentes comunidades. A regiões A e B apresentam cada uma delas cinco habitats, habitados por espécies com diferentes níveis de especialização (largura de nichos).
. Na região A as 10 espécies são generalistas e podem habitar todos os cinco habitats disponíveis. Na região B, as espécies são especialistas e só podem habitar num único habitat. Por conseguinte, toda a região poderá conter 50 espécies já que em cada habitat uma comunidade específica será encontrada. O exemplo ilustra os dois tipos de diversidade: a diversidade alfa ou local e a diversidade beta ou regional. As regiões A e B têm a mesma diversidade alfa, mas a região B tem uma diversidade beta maior. A diversidade alfa depende da capacidade do habitat em suportar um certo número de espécies o que é função do grau de competição entre as populações das diferentes espécies. A diversidade regional depende do grau de substituição de umas espécies pelas outras em diferentes habitats. Tabela 1: Dados hipotéticos de riqueza em espécies em duas regiões com diferentes comunidades. A regiões A e B apresentam cada uma delas cinco habitats, habitados por espécies com diferentes níveis de especialização (largura de nichos).")
6
Os modelos de distribuições das abundâncias das espécies
Os diferentes modelos de curvas de espécie-abundância são consequências das inúmeras tentativas de isolar padrões nesses de abundância das espécies em uma comunidade. O objetivo central destes modelos é achar uma distribuição de probabilidades com um pequeno número de parâmetros (1-3) que variam de uma comunidade para outra e que possam satisfatoriamente explicar e prever os padrões de diversidade existentes na natureza. Quando um ecologista considera o número de espécies numa pequena área com uniformidade de habitat, o grau de especialização em relação aos recursos dentro deste habitat é importante. Quando ele considera o número de espécies numa região maior, o grau de especialização em respeito ao habitat deve ser adicionalmente considerado (ver tabela acima).
 que variam de uma comunidade para outra e que possam satisfatoriamente explicar e prever os padrões de diversidade existentes na natureza. Quando um ecologista considera o número de espécies numa pequena área com uniformidade de habitat, o grau de especialização em relação aos recursos dentro deste habitat é importante. Quando ele considera o número de espécies numa região maior, o grau de especialização em respeito ao habitat deve ser adicionalmente considerado (ver tabela acima).")
7
Distribuições estatísticas
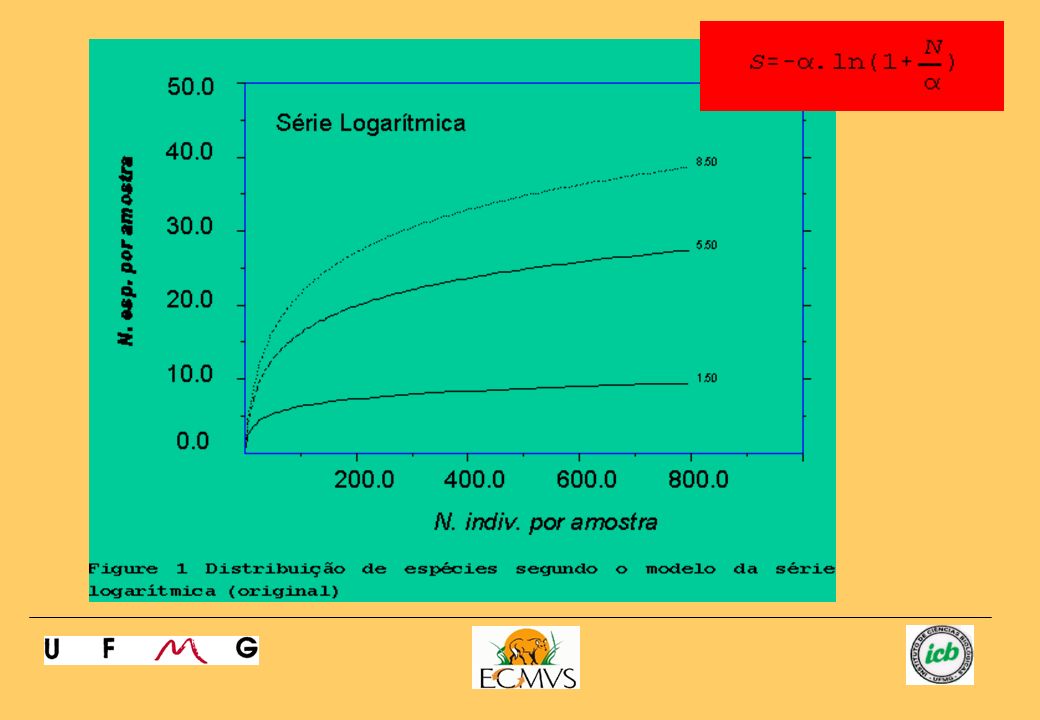
Uma característica encontrada em várias comunidades é a existência de poucas espécies com elevadas abundâncias e muitas espécies com diminutas abundâncias. Neste sentido, alguns modelos estatísticos foram propostos para estudar a comunidade a partir das relações abundância de indivíduos por espécies. Os modelos dados a seguir são os mais citados. 1) Série Logarítimica (Fisher, Cobert & Williams, 1943): A série logarítmica de Fisher é um modelo que tem sido aplicado com sucesso em comunidades relativamente simples, governadas por um único recurso. Ela é definida pelo gráfico e pela equação a seguir:
 Série Logarítimica (Fisher, Cobert & Williams, 1943): A série logarítmica de Fisher é um modelo que tem sido aplicado com sucesso em comunidades relativamente simples, governadas por um único recurso. Ela é definida pelo gráfico e pela equação a seguir:")
9
A figura acima ilustra três comunidades com alfas variando entre 1
A figura acima ilustra três comunidades com alfas variando entre 1.5 e 8.5. Observe que o número final de espécies é dependente do valor de alfa. O parâmetro X não é afetado pelo esforço amostral. O parâmetro alfa também não é afetado pelo esforço amostral e é propriedade intrínseca da comunidade sendo proporcional a S. Na realidade, o parâmetro alfa pode ser tratado como um índice de diversidade. Se S e N são conhecidos, pode-se então solucionar um sistema de equações para alfa e X. O parâmetro deste modelo é o alfa que pode ser obtido pela solução das equações abaixo para alfa e X: onde: S: número de espécies por amostra N: número de indivíduos por amostra alfa: índice de diversidade X: incógnita.

10
O número de espécies com r indivíduos é dado pela fórmula
onde: nr: número de espécies com r indivíduos O modelo de Fisher não funcionou para todas as comunidades por ele testadas. Com esta distribuição, é possível estimar S*, ou seja, o número real de espécies da comunidade.

11
2) Distribuição log-normal (Preston, 1948):
Preston sugeriu expressar o eixo das abscissas (onde estão as abundâncias específicas) numa escala geométrica. Quando esta conversão é feita, a abundância relativa leva a forma de "sino", típica da distribuição normal. Pelo fato do eixo dos x ser expresso em escala logarítmica (por convenção na base 2, mas outras bases numéricas podem igualmente serem usadas), a distribuição passa a ser chamada log-normal. Esta distribuição pode ser definida pela fórmula: onde: yR: número de espécies que ocorrem na r-ésima oitava yo: número de espécies da classe modal (pico do sino) a: constante de dispersão a = sqrt (2.s²) oitava : R = log2 N/No e =
 numa escala geométrica. Quando esta conversão é feita, a abundância relativa leva a forma de sino , típica da distribuição normal. Pelo fato do eixo dos x ser expresso em escala logarítmica (por convenção na base 2, mas outras bases numéricas podem igualmente serem usadas), a distribuição passa a ser chamada log-normal. Esta distribuição pode ser definida pela fórmula: onde: yR: número de espécies que ocorrem na r-ésima oitava. yo: número de espécies da classe modal (pico do sino) a: constante de dispersão a = sqrt (2.s²) oitava : R = log2 N/No. e =")
12
As oitavas nada mais são do que a expressão geométrica das abundâncias dos indivíduos (dados logaritimizados): R = 0 log2 (N/N) R = 1 log2 (2N/N) R = 2 log2 (4N/N) R = 3 log2 (8N/N) R = 4 log2 (16N/N) A oitava Rn refere-se à oitava da classe modal, isto é aquela classe onde ocorre o maior número de espécies. O modelo log-normal de Preston explica um grande número de comunidades. O esforço amostral normalmente "força" a curva para a direita, mas não altera a sua forma. Desde que nós não podemos capturar toda a comunidade, haverá sempre uma parte da curva (espécies raras) que não é representada pela curva. Em consequência, a curva resultante é uma curva truncada assim como a log-série. Preston (1962) demonstrou que as curvas log-normal para a maioria das comunidades biológicas, têm uma configuração especial que ele chamou de distribuição canônica, ou seja, os parâmetros a e yo são interrelacionados (curva canônica). Isto significa que em se sabendo o número de espécies da comunidade pode se especificar todos os outros parâmetros da curva. De posse do parâmetro a, a diversidade pode ser prevista apenas sabendo o número total de espécies se a comunidade se enquadrar no modelo log-normal canônico.
 R = 1 log2 (2N/N) R = 2 log2 (4N/N) R = 3 log2 (8N/N) R = 4 log2 (16N/N) A oitava Rn refere-se à oitava da classe modal, isto é aquela classe onde ocorre o maior número de espécies. O modelo log-normal de Preston explica um grande número de comunidades. O esforço amostral normalmente força a curva para a direita, mas não altera a sua forma. Desde que nós não podemos capturar toda a comunidade, haverá sempre uma parte da curva (espécies raras) que não é representada pela curva. Em consequência, a curva resultante é uma curva truncada assim como a log-série. Preston (1962) demonstrou que as curvas log-normal para a maioria das comunidades biológicas, têm uma configuração especial que ele chamou de distribuição canônica, ou seja, os parâmetros a e yo são interrelacionados (curva canônica). Isto significa que em se sabendo o número de espécies da comunidade pode se especificar todos os outros parâmetros da curva. De posse do parâmetro a, a diversidade pode ser prevista apenas sabendo o número total de espécies se a comunidade se enquadrar no modelo log-normal canônico.")
13
A figura 2 ilustra o modelo log-normal de Preston para três comunidades hipotéticas. A altura do pico do sino depende fundamentalmente de So, ou seja o número de espécies da classe modal (Pinto-Coelho, 2000).
..")
14
A curva log-normal permite estimar o total de espécies, incluindo as espécies ainda não coletadas.
onde: Rn: oitava onde ocorre o número total de indivíduos é máximo Rmax: oitava onde a espécies mais abundante se situa. Para a maioria dos casos vale a relação a seguir:

15
Há muitas evidências de campo indicando que o modelo log-normal se aplica em várias situações:
mariposas na Inglaterra. algas fitoplantônicas no mar Mediterrâneo. serpentes do Panamá. pássaros de New York (USA). O modelo log-normal tem algumas limitações. A primeira delas reside no fato de que não há justificativas biológicas para a lei da curva log-normal. Outro ponto importante liga-se ao fato de que a distribuição log-normal (canônica) assume o estado de equilíbrio estável (stready state).
. O modelo log-normal tem algumas limitações. A primeira delas reside no fato de que não há justificativas biológicas para a lei da curva log-normal. Outro ponto importante liga-se ao fato de que a distribuição log-normal (canônica) assume o estado de equilíbrio estável (stready state).")
16
3) O modelo broken-stick de MacArthur
Este modelo está baseado em interações competitivas entre as espécies de uma comunidade. Os padrões até então conhecidos dependem em grande parte do tipo de organismo coletado. O modelo de MacArthur baseia-se na organização biológica da comunidade. A abundância de cada espécie é determinada pela partição randômica dos recursos distribuídos ao longo de um contínuo de diferentes tipos de recursos. Os recursos são considerados como se estivessem distribuídos ao longo de um bastão (stick em inglês). O esquema a seguir ilustra uma partição aleatória de um recurso (uma dimensão ilustrada pelo segmento ou bastão horizontal) onde foram realizadas cinco partições aleatórias, gerando segmentos ocupados por seis diferentes espécies. Cada segmento corresponderia ao nicho explorado por cada espécie ao longo do recurso (bastão) considerado.
. O esquema a seguir ilustra uma partição aleatória de um recurso (uma dimensão ilustrada pelo segmento ou bastão horizontal) onde foram realizadas cinco partições aleatórias, gerando segmentos ocupados por seis diferentes espécies. Cada segmento corresponderia ao nicho explorado por cada espécie ao longo do recurso (bastão) considerado.")
17
Figura – Partição randômica do pool de espécies de acordo com a disponibilidade de recursos usados pelas espécies de uma comunidade, segundo o modelo do broken stick. Observar que o modelo exige necessariamente que a comunidade esteja saturada em espécies, ou seja, que todos os nichos estejam ocupados (Pinto-Coelho, 2000).
..")
18
O comprimento dos seguimentos é proporcional à abundância relativa das espécies. Se os segmentos são arranjados proporcionalmente numa escala logarítmica decrescente, então a distribuição dos segmentos será aproximadamente linear. O modelo broken stick pode ser descrito pela seguinte fórmula: onde: Na/N : número de indivíduos da espécie a S: número total de espécies a: a-ésima espécie

19
Figura – Distribuição das abundâncias relativas das espécies de três comunidades hipotéticas segundo o modelo broken-stick (Pinto-Coelho, 2000). A figura ilustra o modelo de MacArthur para três comunidades hipotéticas. Observe que o número de espécies de cada comunidade é um parâmetro deste modelo e não uma consequência deste como nos modelos da série logarítmica e log-normal. Observar que a medida que cresce o número de organismos a curva se torna proporcionalmente mais inclinada

20
Características biológicas do modelo broken stick:
Os recursos estão distribuídos continuamente ao longo de uma única dimensão. Todos os recursos disponíveis são utilizados e não há sobreposição de recursos (em inglês resource overlap). O número de espécies é uma premissa do modelo e não uma consequência dele (como na distribuição log-normal). A comunidade deve estar em equilíbrio (steady state ) e deve ser saturada em espécies, ou seja, não deve haver nichos ecológicos vagos. As abundâncias relativas são determinadas por partição ao acaso dos recursos entre as espécies. A competição interespecífica exclui o "niche overlap". O modelo de MacArthur foi testado com sucesso em comunidades caracterizadas por indivíduos de grande tamanho, possuindo ciclo de vida longo (pássaros, gastrópodes, predadores, ofiuróides). Espécies de ciclo de vida curto e de pequeno tamanho (artrópodes do solo, nematóides, fitoplâncton) não se enquandram nesta distribuição. MacArthur sugeriu que apenas as espécies que se encontram em equilíbrio estável poderiam se encaixar neste modelo e as espécies oportunistas tais como muitas espécies de insetos e outros pequenos organismos não se encontram em equilíbrio estável. Hoje sabemos que tais suposições são muitas vezes errôneas e que mesmo em comunidades caracterizadas por pequenos organismos pode haver equilíbrio estável (ver, por exemplo, a teoria da competição de Tilman mais adiante no curso).
. O número de espécies é uma premissa do modelo e não uma consequência dele (como na distribuição log-normal). A comunidade deve estar em equilíbrio (steady state ) e deve ser saturada em espécies, ou seja, não deve haver nichos ecológicos vagos. As abundâncias relativas são determinadas por partição ao acaso dos recursos entre as espécies. A competição interespecífica exclui o niche overlap . O modelo de MacArthur foi testado com sucesso em comunidades caracterizadas por indivíduos de grande tamanho, possuindo ciclo de vida longo (pássaros, gastrópodes, predadores, ofiuróides). Espécies de ciclo de vida curto e de pequeno tamanho (artrópodes do solo, nematóides, fitoplâncton) não se enquandram nesta distribuição. MacArthur sugeriu que apenas as espécies que se encontram em equilíbrio estável poderiam se encaixar neste modelo e as espécies oportunistas tais como muitas espécies de insetos e outros pequenos organismos não se encontram em equilíbrio estável. Hoje sabemos que tais suposições são muitas vezes errôneas e que mesmo em comunidades caracterizadas por pequenos organismos pode haver equilíbrio estável (ver, por exemplo, a teoria da competição de Tilman mais adiante no curso).")
21
Figura - Curvas hipotéticas dos principais modelos vistos acima
Figura - Curvas hipotéticas dos principais modelos vistos acima. As curvas do esquema à direita foram obtidas com dados de campo. Esquema modificado de Magurran (1996), pag. 14. Os dados ilustrados nessa figura vêm de um estudo clássico publicado por Whittaker (1970).
, pag. 14. Os dados ilustrados nessa figura vêm de um estudo clássico publicado por Whittaker (1970).")
Apresentações semelhantes