Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Usando o Código Genético
Capítulo 7 Usando o Código Genético

2
7.1 - Introdução Códons – trios de nucleotídeos organizados do 5’ para o 3’. Correspondem à seqüência de aminoácidos do N-terminal para o C-terminal Estudo do código: Uso de ácido poliuridílico (poliU) polifenilalanina Uso de um trinucleotídeo, que se liga ao ribossomo e a um tRNA.
 polifenilalanina. Uso de um trinucleotídeo, que se liga ao ribossomo e a um tRNA.")
3
7.1 - Introdução Há mais códons (61) do que aminoácidos Códons representando o mesmo ou aminoácidos similares tendem a ter seqüência similar Normalmente a terceira base não é significante. Algumas vezes distingue-se purina de pirimidina.

4
Reações adicionais são permitidas na terceira base
7.1 - Introdução O pareamento dos trinucleotídeos é potencializado pelo ambiente do sítio A ribossomal Reações adicionais são permitidas na terceira base Um mesmo tRNA pode reconhecer mais de um cédon

5
Aminoácidos similares
7.1 - Introdução Aminoácidos similares Códons similares Efeitos de mutações são diminuídos

6
7.1 - Introdução Estabelecimento no começo da evolução Universalidade do código Pequeno número de códons representava pequeno número de aminoácidos Códons mais específicos e maior número de aminoácidos Há exceções para o código universal

7
7.2 – Reconhecimento códon-anticódon
Há sete pares de códons onde o significado é o mesmo independente de qual pirimidina está na terceira posição Há 8 famílias de códons onde códons que tenham as duas primeiras bases têm o mesmo significado. Há três casos em que uma base na terceira posição dá um significado único para um códon: AUG – metionina UGG – triptofano UGA - terminação Há cinco pares de códons a purina na terceira posição não altera o aminoácido

8
7.2 – Reconhecimento códon-anticódon
1º 2º 3º Códon ’ C A U ’ Anticódon ’ G U A ’ 3º 2º 1º

9
7.2 – Reconhecimento códon-anticódon
A base na primeira posição do anticódon pode parear com mais de uma base Normalmente um tRNA reconhece mais de um códon Isso é chamado de wobbling, e ocorre porque a conformação do arco do anticódon dá flexibilidade à primeira base do anticódon

10
7.2 – Reconhecimento códon-anticódon
É possível distinguir códons únicos apenas quando G e U estão na terceira posição

11
7.3 – tRNAs são feitos a partir de precursores maiores
O CCA é adicionado como um processo meramente enzimático Quando um tRNA não é propriamente processado ele é degradado por um sistema de controe de qualidade. tRNAs são sintetizados como cadeias precursoras com material adicional nos dois lados O fim 5’ é gerado pela clivagem de uma enzima chamada ribonuclease P O fim 3’ começa a ser degradado por uma endonuclease que cliva o precursor. A essa seguem várias exonucleases que degradam no sentido 3’ – 5’.

12
7.4 tRNAs contém bases modificadas
tRNAs contém mais de 50 bases modificadas. Modificações a partir das bases A, G, C, U. mRNAs e rRNAs também sofrem modificações de base, mas as alterações dos RNAs transportadores são mais acentuadas. Modificações: metilação, reestruturação do anel...

13
7.4 tRNAs contém bases modificadas
Regiões com bases modificadas: resíduos D braço D ψ seqüência TψC Outras modificações específicas para diferentes grupos: bactérias, leveduras, mamíferos, etc.

14
7.4 tRNAs contém bases modificadas
As enzimas modificadoras de tRNA variam amplamente em tipos e especificidade dentre os diferentes grupos de organismo. aparatos enzimáticos diferentes conjuntos de tRNAs diferentes

15
7.5 Bases modificadas afetam o pareamento anticodon-codon
Modificações de base em quaisquer partes do tRNA afetam sua especificidade. Quando as bases do anticodon são modificadas, tem-se novos padrões de pareamento entre as diferentes bases criadas e as bases A, C, U e G.

16
7.5 Bases modificadas afetam o pareamento anticodon-codon
Inosina(I) pode ser pareada com A, C e U Assim, a existência de I no anticodon de um tRNA permite o reconhecimento de codons de aminoácidos diferentes. Degeneração do código genético
 pode ser pareada com A, C e U. Assim, a existência de I no anticodon de um tRNA permite o reconhecimento de codons de aminoácidos diferentes. Degeneração do código genético.")
17
7.5 Bases modificadas afetam o pareamento anticodon-codon
Padrões de modificação: Inosina, geralmente, é originada de Adenina. A primeira Uridina do anticodon, geralmente, tem suas propriedades de pareamento alteradas.

18
7.5 Bases modificadas afetam o pareamento anticodon-codon
Modificações criam leituras preferenciais de alguns codons Codons utilizados freqüentemente tendem a serem lidos de modo mais eficiente

19
7.5 Bases modificadas afetam o pareamento anticodon-codon
Padrões de modificação diferentes diferentes famílias de tRNAs dentre os organismos.

20
LGCM URLGA 7.6- Há alterações esporáticas no código universal
Tipos de mudanças que podem ocorrer no código genético bacteriano ou no genoma nuclear de eucariotos: 1) Xaa Yaa Ex: CUG Leu Ser Não é comum mudança de um aminoácido para outro. Esta mudança sofre uma forte seleção negativa. 2) Códon de terminação aa Ex: UGA Stop Trp, Cys, Sel Maioria das mudanças são deste tipo, pois não há troca de aminoácido. É necessário modificações no tRNA e nos fatores de terminação para que essa mudança possa acontecer. LGCM URLGA
 Xaa Yaa. Ex: CUG Leu Ser. Não é comum mudança de um aminoácido para outro. Esta mudança sofre uma forte seleção negativa. 2) Códon de terminação aa. Ex: UGA Stop Trp, Cys, Sel. Maioria das mudanças são deste tipo, pois não há troca de aminoácido. É necessário modificações no tRNA e nos fatores de terminação para que essa mudança possa acontecer. LGCM. URLGA.")
21
aa nenhum Ex:AGA Arg nenhum Todas as mudanças são esporáticas. Fig. 7.10

22
Devido as mitocôndrias sintetizarem um pequeno número de proteínas mudanças no seu código genético não seriam fortemente selecionadas. Fig.7.11 A variedade de mudanças encontradas nas mitocôndrias de diferentes espécies sugerem que essas ocorreram separadamente e não em um descendente comum de um código mitocondrial ancestral.

23
7.7 Novos aminoácidos podem ser inseridos no lugar de alguns códons de terminação
1- UGA seleno-cisteínas Ocorrência: procariotos e eucariotos Condições para ocorrer: Presença de seleno-cys-tRNA Estrutura em forma de loop após o códon UGA Fig.7.12 2- UAG pyrrolysine Ocorrência: archaea e bactérias

24
7.8- tRNA são modificados com aminoácido por sintetases
Fig. 7.13 1º aa reage com ATP, formando aminoacil-AMP e liberando pirofosfato 2° aa ativado é transferido para o tRNA, liberando AMP Há 20 tipos de tRNA

25
7.9 Aminoacyl.tRNA synthetases fall into two groups
Grande variedade de enzimas: Subunidades de kD; Monoméricas, diméricas ou tetraméricas. Separação em dois grupos: De acordo com a estrutura do domínio que contém o sítio ativo Domínio catalítico: sítios de ligação para o ATP e aminoácido; Interrompido por um domínio de ligação à hélice aceptora do tRNA Domínio de ligação a região do anticódon do tRNA; Enzimas multiméricas possuem o domíno de formação do oligômero.

26
Variável (depende da enzima)
N.terminal C-terminal Fig – Aminoacil-tRNA sintetase Variável (depende da enzima) Folhas- e -hélices.
 Folhas- e -hélices.")
27
Fig. 7.15 – Aminoacil-tRNA sintetase (estrutura cristalina)
Especula-se que as sintetases ligam-se aos tRNAs pela lateral em L da molécula, principalmente pelas extremidades e que a maior parte da sequência do tRNA não é reconhecida pela enzima.

28
Ligações de hidrogênio
(proteína-tRNA) Fig – tRNA sintetase classe I. Fig – tRNA sintetase classe II. O contato com a enzima altera a estrutura do tRNA
 Fig – tRNA sintetase classe I. Fig – tRNA sintetase classe II. O contato com a enzima altera a estrutura do tRNA.")
29
7.10 Synthetases use proofreading to improve accuracy
Cada sintetase deve ser capaz de distinguir 1 entre 20 aminoácidos e associar-se a 1 tRNA (1-3) em um grupo de cerca de 100 tRNAs. É especialmente difícil distinguir aminoácidos que diferem entre si apenas pelo tamanho da cadeia de carbono A discriminação entre tRNAs é mais fácil já que eles oferecem uma maior superfície de contato. A especificidade de reconhecimento tanto de aminoácidos quanto de tRNAs é controlada pelas aminoacil-tRNA sintetases por reações de prova de leitura que revertem as reações de catálise se um componente incorreto for incorporado.
 em um grupo de cerca de 100 tRNAs. É especialmente difícil distinguir aminoácidos que diferem entre si apenas pelo tamanho da cadeia de carbono. A discriminação entre tRNAs é mais fácil já que eles oferecem uma maior superfície de contato. A especificidade de reconhecimento tanto de aminoácidos quanto de tRNAs é controlada pelas aminoacil-tRNA sintetases por reações de prova de leitura que revertem as reações de catálise se um componente incorreto for incorporado.")
30
As reações de ligação de aminoácidos e tRNAs ocorrem em vários estágios, sendo que os passos evoluem somente se os substratos corretos estiverem ligados à enzima. Como? tRNA se liga ao sítio ativo da enzima tRNA correto tRNA incorreto Mudança conformacional Aminoacilação Fig –Reconhecimento do tRNA pela sintetase.

31
A especificidade por aminoácidos varia de acordo com a enzima.
Algumas enzimas são altamente específicas para a ativação dos aminoácidos enquanto outras não. Quando a sintetase liga um aminoácido incorreto, a revisão requer a ligação do tRNA correto. Pode ocorrer uma mudança conformacional que causa a hidrólise do aminoacil-adenilato incorreto ou então uma transferência do aminoácido para o tRNA, seguido de hidrólise. Fig –Reconhecimento do tRNA pela sintetase.

32
Discriminação entre aminoácidos dependente de tRNA em E. coli
Exemplo: Discriminação entre aminoácidos dependente de tRNA em E. coli Ile-tRNA sintetase pode ligar valina com AMP, mas hidrolisa o valil-adenilato quando tRNAIle é ligado. A taxa de erro global depende da especificidade de passos individuais. 1,5 x 10-5 Taxa bem menor que a de substituição de valina por isoleucina. Isto significa que erros de ligação de aminoácidos são responsáveis por uma fração muito pequena dos erros que na verdade ocorrem durante a síntese protéica.

33
Ile-tRNA sintetase usa o tamanho como forma de discriminação entre os aminoácidos.
Sítio de hidrólise Sítio de ativação Fig Sítios ativos da Ile-tRNA sintetase. O aminoácido é tranportado do sítio de ativação para o de hidrólise da Ile-tRNA sintetase por uma mudança de conformação do tRNA. Fig – Ile-tRNA sintetase.

34
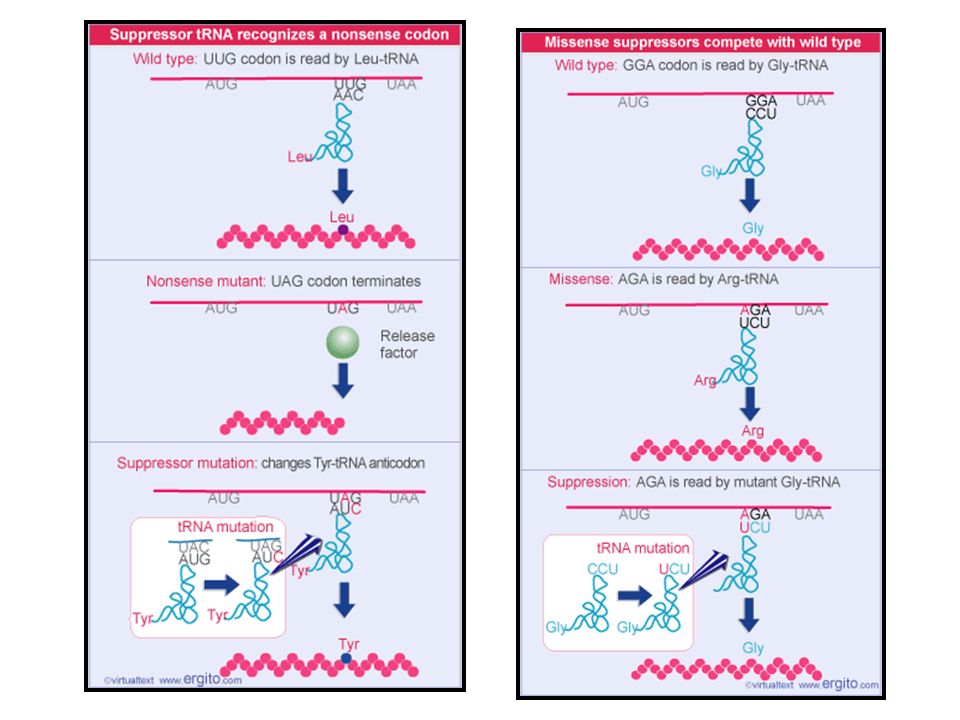
Reconhecendo-o como o códon original
tRNAs supressores Possuem uma mutação no anticódon que altera o códon ao qual esse tRNA responde. tRNAs supressores revertem o efeito de códons mutados Reconhecendo-o como o códon original Restaurando a função protéica

35
tRNAs supressores Nonsense Missense

37
7.12 – Existem supressores nonsense para cada códon de terminação
Os supressores nonsense são divididos em 3 classes, uma para cada tipo de códon de terminação. Supressores âmbar Reconhecem UAG Supressor UGA Seqüência do anticódon não muda Mutação permite o pareamento não usual de C com A Supressores ocre Reconhecem UAA ou UAG

38
O pareamento códon-anticódon tanto de tRNAs selvagens quanto de mutantes não pode ser predito inteiramente da seqüência tripla relevante, mas é influenciado por outras características da molécula.

39
Aminoácido particular
7.13 – Supressores competem com tRNA selvagem Célula selvagem Célula com mutação supressora Códon mutante Códon tRNA supressor Significado usual Aminoácido particular Sinal de terminação Competição

40
Extensão dessa competição vai influenciar a eficiência da supressão
Eficiência de um supressor vai depender: - Afinidade entre seu anticodon e o codon alvo; - Concentração na célula A eficiência de leitura de um códon é influenciada pela sua localização Bases vizinhas ao códon podem mudar a freqüência com a qual um códon é reconhecido por um tRNA particular

41
Supressão nonsnese de terminações naturais
Proteínas alteradas

42
Supressores nonsense serão mais eficientes, quando o codon de terminação reconhecido por ele for relativamente infreqüente no sistema em questão Supressores âmbar em E. coli – códon (UAG) pouco frequente – 10-50% de eficiência Supressores ocre – códon (UAA) usado mais frequentemente – eficiência abaixo de 10% UGA – lido pelo Trp-tRNA com freqüência de 1 a 3% - usado mais comumente que o códon âmbar nos sistemas bacterianos
 pouco frequente – 10-50% de eficiência. Supressores ocre – códon (UAA) usado mais frequentemente – eficiência abaixo de 10% UGA – lido pelo Trp-tRNA com freqüência de 1 a 3% - usado mais comumente que o códon âmbar nos sistemas bacterianos.")
43
Supressores missense Dilema para a célula: suprimir o que é um códon mutante em um lugar e não alterar extensivamente seu significado normal em outros lugares Mutações de uma cópia de um grupo redundante de tRNA Não elimina o reconhecimento dos códons usuais

44
7.14 – O ribossomo influencia a precisão da tradução
A falta de variações detectáveis quando a sequência de uma proteína é analisada demonstra que a síntese proteica é bem precisa. Os poucos erros ocorrem como substituições de um aminoácido por outro. Existem dois estágios na síntese proteica onde erros podem aparecer: 1- ligação do t-RNA com o aminoácido correto 2- reconhecimento códon-anticódon

45
Existem dois modelos básicos para explicar como o ribossomo discrimina entre pareamento correto ou incorreto entre aminoacil-tRNA: 1- A estrutura do ribossomo reconhece aminoacil-tRNAs que paream corretamente. Ou seja, o pareamento correto leva a pequenas mudanças conformacionais que possibilitam a tradução. 2- Existiria dois estágios nesse processo, assim o aminoacil-tRNA teria várias oportunidades de se desligar do ribossomo. Um pareamento errôneo impediria a passagem de um estado para o outro.

46
O modelo atual incorpora elementos dos dois outros modelos propostos:

47
7.15 – Recoding changes codon meanings
A fase de leitura é geralmente invariável. Começa em AUG e continua em triplets até o códon de terminação. A leitura não reconhece senso. Ou seja, a inserção ou a deleção leva a um “frameshift”. Existem algumas exceções dos padrões de tradução que possibilitam que uma janela de leitura com interrupções – códon sem sentido ou frameshift – possa ser traduzida em uma proteína inteira. Mundando o sentido de um códon permite-se colocar um aminoácido em um códon de terminação, ou inserir um aminoácido no lugar de outro.

48
Uma mutação em um tRNA (geralmente no anticódon) pode suprimir o sentido original do códon. Em um caso especial, um tRNA específico é acoplado por um fator de elongação para reconhecer um códon de terminação adjacente a um loop em grampo

49
7.16 Frameshift ocorre em seqüências “escorregadias”
Frameshifting está associado com tRNAs específicos: Supressores de tRNAs mutantes reconhecem um códon de 4 bases certas seqüências permitem o tRNA mover-se ao longo do mRNA no sítio A Mutações por frameshift ocorrem pela inserção ou deleção de uma base e podem ser suprimidas restaurando a fase de leitura original O tipo mais simples de supressor de frameshift corrige a fase de leitura quando houve uma inserção em um trecho de bases repetidas

50
Em fagos e vírus, ocorrem situações em que frameshifting é um evento normal
esses eventos podem causar a terminação ou continuação da síntese protéica Ex. tradução em retrovírus

51
Frameshifting programado ocorre com freqüências 100-1000 x maiores
seqüências escorregadias permitem o aminoacil-tRNA mover-se 1 base para frente ou para trás o ribossomo é atrasado para permitir o rearranjo

52
7.17 Ultrapassagem envolve movimentação do ribossomo
Algumas seqüências promovem evento de ultrapassagem evento mais raro, apenas 3 exemplos (Ex.: gene60 do fago T4) códons idênticos nas extremidades da seqüência omitida
 códons idênticos nas extremidades da seqüência omitida.")
53
Também depende de pausa do ribossomo
maior probabilidade de dissociação do peptidil-tRNA quando há atraso na entrada de aminoacil-tRNA Gene 60 do fago T4: estrutura do mRNA reduz eficiência da terminação

Apresentações semelhantes