Carregar apresentação
A apresentação está carregando. Por favor, espere

1
FIGURA 4. 1 Síntese semiconservativa do DNA
FIGURA 4.1 Síntese semiconservativa do DNA. As duas fitas parentais se separam e cada uma é molde para a síntese de uma fita filha complementar. Portanto, os dois duplex filhos têm cada um uma fita parental e uma fita recém-sintetizada.

2
FIGURA 4.5 Separação das fitas parentais em uma forquilha de replicação. Síntese de DNA ocorre em uma estrutura chamada forquilha de replicação, na qual as fitas parentais são separadas. Uma helicase é necessária para separar fitas parentais e permitir que a forquilha progrida. Proteínas que se ligam a DNA de fita simples (single-stranded DNA-binding proteins – SSB) são necessárias para manter as fitas separadas. As fitas parentais enrolam-se uma em torno da outra mais ou menos uma vez a cada 10,5 pb, e devem se desenrolar para se separarem; isso gera problemas topológicos. Note que uma das fitas em crescimento está orientada com sua extremidade 3’ voltada para a forquilha, enquanto a outra está orientada com sua extremidade 5’ voltada para a forquilha.
 são necessárias para manter as fitas separadas. As fitas parentais enrolam-se uma em torno da outra mais ou menos uma vez a cada 10,5 pb, e devem. se desenrolar para se separarem; isso gera problemas topológicos. Note que uma das fitas em crescimento está orientada com sua extremidade 3’ voltada para a forquilha, enquanto a outra está orientada com sua extremidade 5’ voltada para a forquilha.")
3
FIGURA 4.6 Resolvendo o problema de polaridade: síntese semidescontínua de DNA. Ambas as fitas são elongadas em suas extremidades 3’. A que tem sua extremidade 3’ voltada para a forquilha pode crescer continuamente; é chamada fita contínua ou líder. Como nenhuma DNA polimerase pode adicionar nucleotídeos à extremidade 5’ de uma cadeia em crescimento, a fita com sua extremidade 5’ voltada para a forquilha é feita como uma série de pequenos pedaços chamados fragmentos de Okazaki, cada um sintetizado na direção de 5’ para 3’, e depois unidos. Esta é chamada de fita descontínua, retrógrada ou tardia.

4
FIGURA 4. 7Síntese de DNA em uma forquilha de replicação
FIGURA 4.7Síntese de DNA em uma forquilha de replicação. (a) A fita contínua é elongada em sua extremidade 3’ por adições repetidas de nucleotídeos, como mostra a Figura 4.2. Um fragmento de Okazaki feito previamente, com um pequeno primer de RNA, é mostrado do outro lado. (b) Usando a fita parental como molde, primase sintetiza um primerde RNA (preto). (c) DNA polimerase estende covalentemente a extremidade 3’ do primerde RNA, incorporando desoxirribonucleotídeos (cinza médio), como mostra a Figura 4.2. (d) Quando o fragmento de Okazaki aproxima-se do fragmento sintetizado anteriormente, o primer de RNA do fragmento mais antigo é removido e a falha é preenchida por uma DNA polimerase que elonga o fragmento de Okazaki mais novo. Enquanto isso, a forquilha abre mais e primase sintetizou um novo primer. (e) Quando a falha foi preenchida e apenas uma quebra permanece, uma DNA ligase sela os dois fragmentos (dentro do círculo). Esse processo será repetido com o novo primer elongado, o primer antigo removido, a falha preenchida e a quebra selada.
 A fita contínua é elongada em sua extremidade 3’ por adições repetidas de nucleotídeos, como mostra a Figura 4.2. Um fragmento de Okazaki feito previamente, com um pequeno primer de RNA, é mostrado do outro lado. (b) Usando a fita parental como molde, primase sintetiza um primerde RNA (preto). (c) DNA polimerase estende covalentemente a extremidade 3’ do primerde RNA, incorporando desoxirribonucleotídeos (cinza médio), como mostra a Figura 4.2. (d) Quando o fragmento de Okazaki aproxima-se do fragmento sintetizado anteriormente, o primer de RNA do fragmento mais antigo é removido e a falha é preenchida por uma DNA polimerase que elonga o fragmento de Okazaki mais novo. Enquanto isso, a forquilha abre mais e primase sintetizou um novo primer. (e) Quando a falha foi preenchida e apenas uma quebra permanece, uma DNA ligase sela os dois fragmentos (dentro do círculo). Esse processo será repetido com o novo primer elongado, o primer antigo removido, a falha preenchida e a quebra selada.")
5
FIGURA 4. 10Braçadeiras corrediças
FIGURA 4.10Braçadeiras corrediças. (a) Uma proteína de instalação da braçadeira (clamp-loading) se liga ao DNA. (b) O instalador de braçadeira organiza a braçadeira corrediça a partir de suas subunidades. (c) DNA polimerase associa-se com a braçadeira organizada e torna-se processiva. (d) Estrutura da braçadeira corrediça em células de mamíferos, um trímero de subunidades PCNA. O DNA passa livremente pelo grande buraco no centro do complexo.
 Uma proteína de instalação da braçadeira (clamp-loading) se liga ao DNA. (b) O instalador de braçadeira organiza a braçadeira corrediça a partir de suas subunidades. (c) DNA polimerase associa-se com a braçadeira organizada e torna-se processiva. (d) Estrutura da braçadeira corrediça em células de mamíferos, um trímero de subunidades PCNA. O DNA passa livremente pelo grande buraco no centro do complexo.")
6
FIGURA 4. 11 Replicação de ambas as fitas num replissomo
FIGURA 4.11 Replicação de ambas as fitas num replissomo. Pela ligação de duas DNA polimerases juntas e a fita descontínua criando uma alça para que possa passar através do complexo, ambas as fitas podem ser feitas em um só lugar. O DNA passa pelo complexo, girando à medida que passa; o grande complexo protéico não precisa girar ao redor do DNA. FIGURA 4.12 Enzimas da replicação em E.coli. A fita contínua (acima) é elongada por pol III associada a uma braçadeira corrediça. O complexo DNA helicase/primase (DnaB/DnaG) move-se ao longo do molde para a fita tardia (abaixo); faz uma pausa para permitir que a DnaG sintetize um primer de RNA a cada 1000 a 2000 nt. Pol III elonga o primer até que ele alcance o primer do fragmento de Okazaki sintetizado anteriormente. Nesse momento, pol III libera o DNA e pol I se liga à extremidade do fragmento de Okazaki. Pol I remove o primer de RNA, um nucleotídeo de cada vez, usando sua atividade 5’-endonuclease e simultaneamente preenche a falha em um processo chamado “nick translation”. Quando há uma quebra remanescente com desoxirribonucleotídeos em ambos os lados, a DNA ligase pode selar a quebra.
 é elongada por pol III associada a uma braçadeira corrediça. O complexo DNA helicase/primase (DnaB/DnaG) move-se ao longo do molde para a fita tardia (abaixo); faz uma pausa para permitir que a DnaG sintetize um primer de RNA a cada 1000 a 2000 nt. Pol III elonga o primer até que ele alcance o primer do fragmento de Okazaki sintetizado anteriormente. Nesse momento, pol III libera o DNA e pol I se liga à extremidade do fragmento de Okazaki. Pol I remove o primer de RNA, um nucleotídeo de cada vez, usando sua atividade 5’-endonuclease e simultaneamente preenche a falha em um processo chamado nick translation . Quando há uma quebra remanescente com desoxirribonucleotídeos em ambos os lados, a DNA ligase pode selar a quebra.")
7
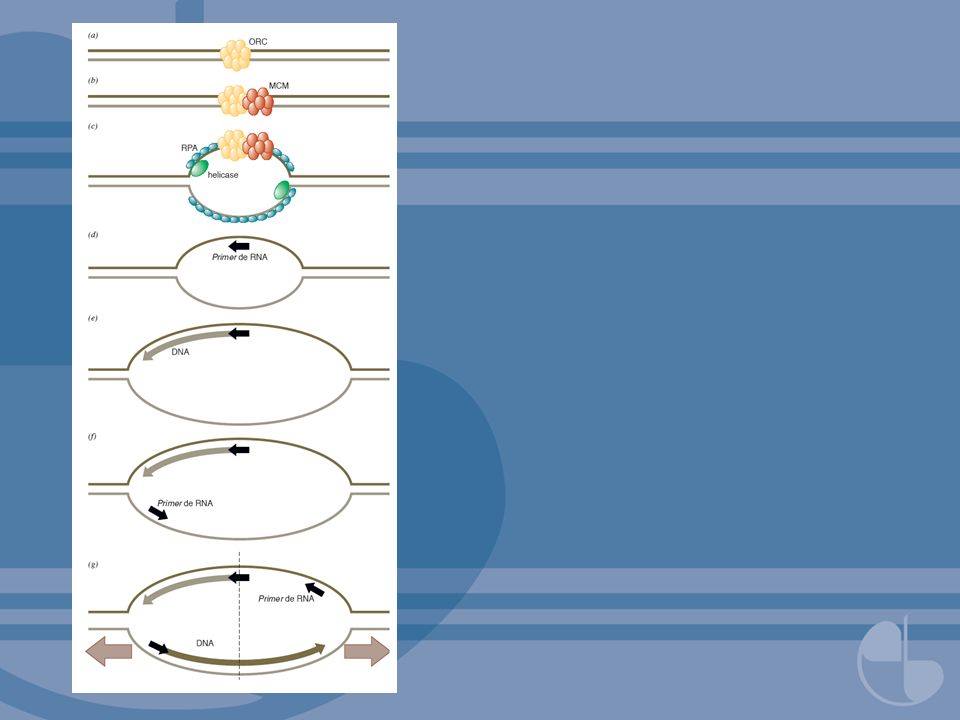
FIGURA 4.13 Enzimas replicando a fita tardia em eucariotos (a fita líder não é mostrada). (a) RPA, uma SSB, liga-se ao molde de fita simples que foi aberto pela progressão da forquilha de replicação. Um fragmento de Okazaki anterior é mostrado à direita, com o primer de RNA em cinza escuro. (b) A atividade da primase no complexo pol α/primase inicia a síntese de um primer de RNA. (c) Depois de o primer de RNA atingir 10 nucleotídeos, a atividade polimerase do complexo pol α/primase assume e elonga o primer com cerca de desoxinucleotídeos. Então, pol α/primase dissocia-se. (d) RFC se liga à extremidade deste fragmento de Okazaki parcialmente completo e catalisa a montagem de uma braçadeira corrediça a partir de três moléculas de PCNA. (e) Pol δ se liga à braçadeira corrediça e elonga o fragmento de Okazaki. (f) À medida que o complexo de replicação aproxima-se do primer de RNA anterior, esse primer é degradado pela ação combinada de RNase H e FEN1. (g) A falha é preenchida pela continuação da elongação do fragmento de Okazaki. (h) A quebra remanescente é selada pela DNA ligase.
 RPA, uma SSB, liga-se ao molde de fita simples que foi aberto pela progressão da forquilha de replicação. Um fragmento de Okazaki anterior é mostrado à direita, com o primer de RNA em cinza escuro. (b) A atividade da primase no complexo pol α/primase inicia a síntese de um primer de RNA. (c) Depois de o primer de RNA atingir 10 nucleotídeos, a atividade polimerase do complexo pol α/primase assume e elonga o primer com cerca de desoxinucleotídeos. Então, pol α/primase dissocia-se. (d) RFC se liga à extremidade deste fragmento de Okazaki parcialmente completo e catalisa a montagem de uma braçadeira corrediça a partir de três moléculas de PCNA. (e) Pol δ se liga à braçadeira corrediça e elonga o fragmento de Okazaki. (f) À medida que o complexo de replicação aproxima-se do primer de RNA anterior, esse primer é degradado pela ação combinada de RNase H e FEN1. (g) A falha é preenchida pela continuação da elongação do fragmento de Okazaki. (h) A quebra remanescente é selada pela DNA ligase..")
8
FIGURA 4.14 Replicons em seqüência e replicação bidirecional em eucariotos. (a) Há múltiplas origens de replicação (ori) organizadas em seqüência ao longo de cada cromossomo eucariótico. No homem, são espaçadas em intervalos de aproximadamente pb. (b) Início ocorre em todas as origens (ori). Grupos adjacentes de oristendem a funcionar juntos. (c) Duas forquilhas de replicação divergentes são estabelecidas em cada ori, de modo que a replicação é bidirecional. As estruturas formadas são chamadas bolhas de replicação. (d) Bolhas de replicação aumentam, à medida que a replicação continua, até que fiquem bem próximas. Nesse estágio, o término da replicação une bolhas adjacentes e separa o DNA parental. Topoisomerase II é essencial para término e segregação dos cromossomos. (e) Os cromossomos duplicados resultantes podem, então, segregar em duas células filhas.
 Há múltiplas origens de replicação (ori) organizadas em seqüência ao longo de cada cromossomo eucariótico. No homem, são espaçadas em intervalos de aproximadamente pb. (b) Início ocorre em todas as origens (ori). Grupos adjacentes de oristendem a funcionar juntos. (c) Duas forquilhas de replicação divergentes são estabelecidas em cada ori, de modo que a replicação é bidirecional. As estruturas formadas são chamadas bolhas de replicação. (d) Bolhas de replicação aumentam, à medida que a replicação continua, até que fiquem bem próximas. Nesse estágio, o término da replicação une bolhas adjacentes e separa o DNA parental. Topoisomerase II é essencial para término e segregação dos cromossomos. (e) Os cromossomos duplicados resultantes podem, então, segregar em duas células filhas..")
10
FIGURA 4.16 Ciclo celular. Em células eucarióticas, a replicação do DNA (fase S) e a mitose (fase M) são separadas por dois intervalos, G1 e G2. O tamanho dos segmentos representa a fração típica do tempo ciclo de divisão celular. FIGURA 4.18 Telomerase. Telomerase é um complexo ribonucleoprotéico com uma fita curta de RNA, como parte integral; catalisa a adição de novas repetições teloméricas de 6-nt à extremidade 3’ de uma cadeia de DNA. O RNA da telomerase pareia parcialmente pelas bases com a repetição telomérica e serve de molde para a reação, enquanto o componente protéico funciona como uma transcriptase reversa, sintetizando DNA usando o RNA como molde. Depois da adição de uma repetição de seis nucleotídeos, a enzima pode se dissociar e se ligar novamente e adicionar novas repetições de 6-nt.

14
FIGURA 4. 23 Lesão no DNA. (a) Deaminação oxidativa converte C em U
FIGURA 4.23 Lesão no DNA. (a) Deaminação oxidativa converte C em U. (b) Sítio AP. Despurinação (ou, menos freqüentemente, despirimidinação) é a clivagem de uma ligação glicosídica entre a posição 1´ do açúcar e a base. Isso deixa um sítio abásico ou AP, mas não quebra o esqueleto açúcar-fosfato. (c) O6-metil guanosina é uma lesão altamente mutagênica. (d) Luz ultravioleta leva à ligação entre piridinas adjacentes ao longo de uma das fitas do DNA. Um dímero de timina cis-syn ciclobutano é mostrado no esquema. (e) Um dímero ciclobutano é mostrado na dupla fita do DNA; o esqueleto é mostrado como um bastão para revelar a distorção das bases em ligação cruzada (circulado em preto) e bases complementares na outra fita.
 Deaminação oxidativa converte C em U. (b) Sítio AP. Despurinação (ou, menos freqüentemente, despirimidinação) é a clivagem de uma ligação glicosídica entre a posição 1´ do açúcar e a base. Isso deixa um sítio abásico ou AP, mas não quebra o esqueleto açúcar-fosfato. (c) O6-metil guanosina é uma lesão altamente mutagênica. (d) Luz ultravioleta leva à ligação entre piridinas adjacentes ao longo de uma das fitas do DNA. Um dímero de timina cis-syn ciclobutano é mostrado no esquema. (e) Um dímero ciclobutano é mostrado na dupla fita do DNA; o esqueleto é mostrado como um bastão para revelar a distorção das bases em ligação cruzada (circulado em preto) e bases complementares na outra fita.")
15
FIGURA 4. 25 Reparo por excisão de base
FIGURA 4.25 Reparo por excisão de base. (a) O ponto danificado (bolinha colocada na fita (a)) é reconhecido por uma DNA glicosilase. (b) A base é removida por clivagem da ligação glicosídica que a conecta com o açúcar desoxiribose; o esqueleto açúcar-fosfato não é quebrado. Isso deixa um sítio AP. (c) O esqueleto açúcar-fosfato é clivado no lado 5’ do sítio abásico por uma AP endonuclease. Há também clivagem no lado 3’ desse sítio para remover o resíduo de açúcar; isso pode ser feito por algumas AP endonucleases ou por uma atividade de AP liase. (d) A falha de um único nucleotídeo é preenchida por uma DNA polimerase. (e) A quebra remanescente é selada por uma DNA ligase.
 O ponto danificado (bolinha colocada na fita (a)) é reconhecido por uma DNA glicosilase. (b) A base é removida por clivagem da ligação glicosídica que a conecta com o açúcar desoxiribose; o esqueleto açúcar-fosfato não é quebrado. Isso deixa um sítio AP. (c) O esqueleto açúcar-fosfato é clivado no lado 5’ do sítio abásico por uma AP endonuclease. Há também clivagem no lado 3’ desse sítio para remover o resíduo de açúcar; isso pode ser feito por algumas AP endonucleases ou por uma atividade de AP liase. (d) A falha de um único nucleotídeo é preenchida por uma DNA polimerase. (e) A quebra remanescente é selada por uma DNA ligase.")
16
FIGURA 4. 26Reparo por excisão do nucleotídeo
FIGURA 4.26Reparo por excisão do nucleotídeo. (a) A base danificada (pequena bolinha nas fitas (a) e (b)) é reconhecida por um complexo de reparo do DNA. (b) O segmento ao redor da base lesada é removido por um complexo enzimático que faz duas quebras na fita danificada, uma de cada lado da lesão. (c) Um oligonucleotídeo (27-29 nt no homem, nt em E.coli) é liberado. (d) Ressíntese: uma DNA polimerase preenche a falha, usando a fita oposta como molde. (e) A DNA ligase sela a quebra remanescente.
 A base danificada (pequena bolinha nas fitas (a) e (b)) é reconhecida por um complexo de reparo do DNA. (b) O segmento ao redor da base lesada é removido por um complexo enzimático que faz duas quebras na fita danificada, uma de cada lado da lesão. (c) Um oligonucleotídeo (27-29 nt no homem, nt em E.coli) é liberado. (d) Ressíntese: uma DNA polimerase preenche a falha, usando a fita oposta como molde. (e) A DNA ligase sela a quebra remanescente.")
17
FIGURA 4. 28 Reparo de pareamento errado em E. coli
FIGURA 4.28 Reparo de pareamento errado em E.coli. (a) Um par errado no DNA recém -replicado. A fita parental está metilada, mas nos primeiros minutos após sua síntese, a nova fita ainda não foi metilada; tal sítio, com apenas uma das fitas metilada, é chamada hemimetilado. (b) MutS e MutL ligam-se ao par errado. ATP é hidrolisado quando desloca uma pequena alça do DNA. (c, f) MutH se liga a um sítio hemimetilado (que poderia estar no lado esquerdo ou direito do par errado). (d, g) A fita não-metilada é quebrada e depois removida, estendendo-se para além do ponto do par errado. Isso pode ocorrer em qualquer direção (d e g). (e, h) A longa falha resultante é preenchida por uma DNA polimerase, adicionando nucleotídeos à extremidade 3’ da fita (preta). (i) A quebra remanescente é selada por uma DNA ligase (fita de (e) é mostrada). (j) Os sítios hemimetilados são metilados (fita de (h) é mostrada).
 Um par errado no DNA recém -replicado. A fita parental está metilada, mas nos primeiros minutos após sua síntese, a nova fita ainda não foi metilada; tal sítio, com apenas uma das fitas metilada, é chamada hemimetilado. (b) MutS e MutL ligam-se ao par errado. ATP é hidrolisado quando desloca uma pequena alça do DNA. (c, f) MutH se liga a um sítio hemimetilado (que poderia estar no lado esquerdo ou direito do par errado). (d, g) A fita não-metilada é quebrada e depois removida, estendendo-se para além do ponto do par errado. Isso pode ocorrer em qualquer direção (d e g). (e, h) A longa falha resultante é preenchida por uma DNA polimerase, adicionando nucleotídeos à extremidade 3’ da fita (preta). (i) A quebra remanescente é selada por uma DNA ligase (fita de (e) é mostrada). (j) Os sítios hemimetilados são metilados (fita de (h) é mostrada).")
18
FIGURA 4. 30Fotorreativação
FIGURA 4.30Fotorreativação. (a) Fotoliase liga-se a um ciclobutano dímero de pirimidina; luz não é necessária para essa etapa. (b) O complexo absorve luz, o que resulta na clivagem das ligações unindo pirimidinas adjacentes. (c) A enzima então se dissocia do DNA
 Fotoliase liga-se a um ciclobutano dímero de pirimidina; luz não é necessária para essa etapa. (b) O complexo absorve luz, o que resulta na clivagem das ligações unindo pirimidinas adjacentes. (c) A enzima então se dissocia do DNA.")
19
FIGURA 4. 31Síntese bypass (translesão)
FIGURA 4.31Síntese bypass (translesão). (a) Lesão no molde geralmente faz parar DNA polimerases replicativas, uma vez que não podem adicionar nucleotídeo oposto a uma lesão não-codificadora ou com distorção na fita. (b) DNA polimerases especializadas, com especificidade relaxada (ver Tabela 4.2) podem inserir nucleotídeos opostos a lesões no molde permitindo, assim, ultrapassar (bypass) a lesão e sintetizar fitas filhas sem falhas. A especificidade relaxada e a falta de revisão dessas polimerases significam que erros (mutações em potencial) são freqüentemente introduzidos. Essas polimerases se dissociam facilmente do DNA, de modo que o comprimento do DNA que sintetizam é geralmente pequeno. FIGURA 4.32Reparo de falha na fita filha. (a) Falhas ficam no DNA recém-replicado, onde forquilhas de replicação param devido a lesões. Como a lesão não tem DNA intacto oposto, não é substrato para reparo por excisão. (b) Recombinação permite que a fita parental isopolar (cinza-escuro) preencha a falha na fita filha (cinza-claro). Isso deixa uma falha na fita parental. (c) A falha na fita parental pode ser preenchida (preto) por uma DNA polimerase, porque há um molde intacto oposto. O resultado é reparo da falha na fita filha, mas a lesão permanece. Note que a lesão agora pode ser reparada por reparo de excisão, porque agora está oposta a uma fita intacta.
. (a) Lesão no molde geralmente faz parar DNA polimerases replicativas, uma vez que não podem adicionar nucleotídeo oposto a uma lesão não-codificadora ou com distorção na fita. (b) DNA polimerases especializadas, com especificidade relaxada (ver Tabela 4.2) podem inserir nucleotídeos opostos a lesões no molde permitindo, assim, ultrapassar (bypass) a lesão e sintetizar fitas filhas sem falhas. A especificidade relaxada e a falta de revisão dessas polimerases significam que erros (mutações em potencial) são freqüentemente introduzidos. Essas polimerases se dissociam facilmente do DNA, de modo que o comprimento do DNA que sintetizam é geralmente pequeno. FIGURA 4.32Reparo de falha na fita filha. (a) Falhas ficam no DNA recém-replicado, onde forquilhas de replicação param devido a lesões. Como a lesão não tem DNA intacto oposto, não é substrato para reparo por excisão. (b) Recombinação permite que a fita parental isopolar (cinza-escuro) preencha a falha na fita filha (cinza-claro). Isso deixa uma falha na fita parental. (c) A falha na fita parental pode ser preenchida (preto) por uma DNA polimerase, porque há um molde intacto oposto. O resultado é reparo da falha na fita filha, mas a lesão permanece. Note que a lesão agora pode ser reparada por reparo de excisão, porque agora está oposta a uma fita intacta.")
20
FIGURA 4. 33 Regulação do reparo do DNA e recuperação em E
FIGURA 4.33 Regulação do reparo do DNA e recuperação em E. coli : a respos-ta SOS. (a) Em células não danificadas, um grupo de operons que constituem o regulon SOS é regulado por um repressor comum, LexA. Esse regulon inclui o próprio gene lexA (auto-regulação) e o gene recA, juntamente com muitos genes que codificam enzimas que agem reparando lesões. Há uma transcrição baixa, constitutiva de muitos desses genes, incluindo lexA e recA, mesmo quando a proteína LexA está presente. (b) Lesões bloqueiam forquilhas de replicação e, assim, deixam DNA fita-simples. RecA se liga a DNA fita-simples e é “ativada”. Nessa forma, ajuda na clivagem de LexA. Os fragmentos clivados de LexA não podem se ligar aos operado-res, de modo que todo o conjunto de operons é induzido. Muito mais RecA é feita, juntamente com outras proteínas, incluindo LexA. Enquanto existirem lesões, RecA permanecerá ativada, LexA continuará a ser clivada para que os operons permaneçam ligados. Quando as lesões são reparadas, RecA é “desativada” e não mais ajuda a clivar LexA, de modo que LexA ativa se acumula e desliga os genes desse regulon.
 Em células não danificadas, um grupo de operons que constituem o regulon SOS é regulado por um repressor comum, LexA. Esse regulon inclui o próprio gene lexA (auto-regulação) e o gene recA, juntamente com muitos genes que codificam enzimas que agem reparando lesões. Há uma transcrição baixa, constitutiva de muitos desses genes, incluindo lexA e recA, mesmo quando a proteína LexA está presente. (b) Lesões bloqueiam forquilhas de replicação e, assim, deixam DNA fita-simples. RecA se liga a DNA fita-simples e é ativada . Nessa forma, ajuda na clivagem de LexA. Os fragmentos clivados de LexA não podem se ligar aos operado-res, de modo que todo o conjunto de operons é induzido. Muito mais RecA é feita, juntamente com outras proteínas, incluindo LexA. Enquanto existirem lesões, RecA permanecerá ativada, LexA continuará a ser clivada para que os operons permaneçam ligados. Quando as lesões são reparadas, RecA é desativada e não mais ajuda a clivar LexA, de modo que LexA ativa se acumula e desliga os genes desse regulon.")
Apresentações semelhantes