Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Biologia dos marcadores moleculares Almir R. Pepato

2
Ácidos Nucléicos PirimidinaPurina

3
IUPAC Código formalizado pela International Union of Pure and Applied Chemistry (IUPAC) em1970.
 em1970.")
4
RNA Nos vírus de RNA (fita simples ou dupla), funciona como material genético Mas também é capaz de produzir configurações espaciais complexas e com isso apresentar capacidade catalítica análoga às enzimas protéicas.
, funciona como material genético Mas também é capaz de produzir configurações espaciais complexas e com isso apresentar capacidade catalítica análoga às enzimas protéicas.")
5
RNA

6
Mundo de RNA, hoje DNA RNAProteínas Armazenamento de informação Assumiram a maior parte das atividades catalíticas e estruturais das células. rRNA, tRNA, mRNA, e muito mais:

7
DNA Polimerase

8
Mutações Bases nitrogenadas anômalas são uma das causas mais comuns de mutações pontuais.

9
Mutações

10
Transcrição

11
Processamento do mRNA O splicing alternativo pode constituir um importante mecanismo de evolução das proteínas, permitindo combinações originais de sítios funcionais.

12
Processamento do mRNA

13
Transcriptase reversa

14
Síntese protéica

15
Código genético Codifica para os 20 aminoácidos e códons de parada.

16
Questões para discussão 1- O DNA é um ácido nucléico. Sua carga elétrica é NEGATIVA, submetido a um campo elétrico, portanto, correrá do pólo NEGATIVO para o POSITIVO.

17
Questões para discussão 2- Transições são substituições entre pirimidinas ou entre purinas. Transversões são substituições de purinas por pirimidinas e vice-versa. Complete a matriz de custos abaixo, onde às transições é atribuído o custo 1 e às transversões valor 2. Na maioria das sequências as transições são mais frequentes que as transversões.

18
Questões para discussão 3- Qual das seguintes mutações afetando códons em uma proteína de um gene nuclear deve ter MENOS impacto sobre a aptidão do organismo? Seq. original: UUU UAU GAG CUU Phe Tyr Glu Leu Mutação 1: UUU UAU GUG CUU Mutação 2: UUU UAA GAG CUU Mutação 3: - UUU AUG AGC UU... Mutação 4: UUC UAC GAG UUG Phe Tyr Val Leu Phe --- --- --- Phe Met Ser... Phe Tyr Glu Leu

19
Marcadores mais utilizados Almir R. Pepato

20
Definição de marcador molecular Uma sequência nucleotídica ou de aminoácidos detectável experimentalmente

21
Genes Ribossomais Metáfase de Serrasalmus serrulatus marcadas para o 18S (Nakayama et al, 2008) Todos os genomas apresentam genes ribossomais (Procariotos, Mitocôndrias, Cloroplastos, Nucleares) Nos Eucariotos há várias cópias agrupadas em diversos cromossomos
 Todos os genomas apresentam genes ribossomais (Procariotos, Mitocôndrias, Cloroplastos, Nucleares) Nos Eucariotos há várias cópias agrupadas em diversos cromossomos")
22
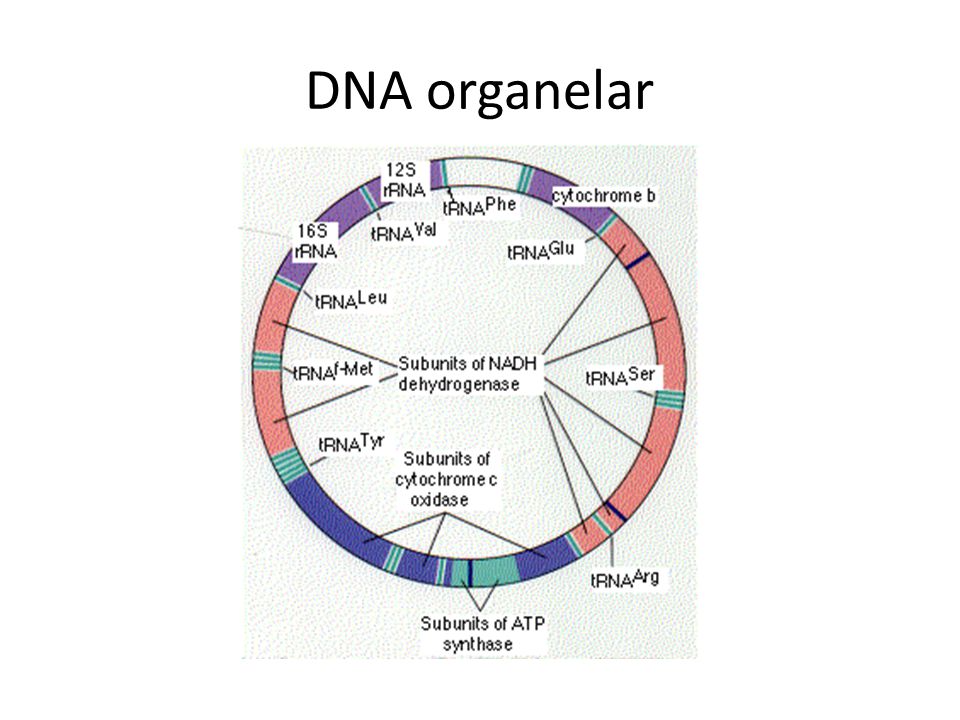
Estrutura dos genes Ribossomais Nas mitocôndrias esses genes são ainda mais compactos: SSU: 12S LSU: 16S

23
Estrutura secundária

25
Vantagens do emprego dos rDNA Primers conservados Múltiplas cópias, evolução concertada Poucos problemas de paralogia É um dos únicos marcadores que pode ser sequenciado em qualquer ser vivo A estrutura secundaria auxília no alinhamento das sequências e fornece outros caracteres.

26
Genes codificantes

27
Dificuldades técnicas: Poucas cópias na amostra reduz a quantidade de substrato para a reação de PCR. Propriedades semelhantes aos dos genes ribossomais

28
Genes codificantes Exemplo de gene com multiplas cópias, com evidências de evolução concertada.

29
Genes codificantes Alternativa para a obtenção de genes de cópia simples: Transcriptase Reversa- PCR (Utilizado também para ESTs, apenas um artigo).
.")
30
ITS e outros introns Os introns são regiões excluídas do mRNA graças ao mecanismo de splicing. Como são regiões que não codificam proteínas estão menos sujeitas à seleção natural estabilizadora e assim acumulam mais substituições. Geralmente empregados para recuperar histórias evolutivas recentes.

31
DNA organelar

33
Fênomeno comum nos genomas mitocôndriais, o viés no emprego de nucleotídeos pode levar a erros nas inferências filogenéticas

34
Mutação Vs Substituição Mutação é um fenômeno químico. Produz novas versões dos genes. Substituição é um fenômeno populacional.

35
Mecanismos que levam à fixação de alelos Deriva gênica: No caso do aparecimento de uma nova mutação, m=1: Considerando uma taxa de mutação μ:

36
Mecanismos que levam à fixação de alelos Modelo Wright-Fisher para descrever a evolução por deriva gênica: Probabilidade de ter a mutação: Probabilidade de não ter a mutação: Isso dá uma distribuição binomial, com a probabilidade de termos n mutantes na geração seguinte de:

37
Mecanismos que levam à fixação de alelos Seleção natural, aptidão média: A probabilidade de que o gene mutante seja transmitido à nova geração é de: Agora basta substituir “a” no modelo de Wright- Fisher.

38
Coalescência Exemplo de um modelo simples: Em uma população em que todos os indivíduos apresentam o mesmo número médio de descendentes a probabilidade de um indivíduos compartilhar a mãe é de: Já a possibilidade de não compartilharem é de:

39
Coalescência A probabilidade de dois indivíduos compartilharem um dos pais a T gerações atrás é de : Ou: O tempo para a coalescência nas nossas condições inverossímeis é 2N.

40
Cenários para a evolução molecular

41
Princípios da genética molecular –Hubby e Lewontin, 1966; Harris, 1966 Revelou um nível de polimorfismo insuspeito.

42
Relógio molecular Dickerson, 1971 Proporcional ao tempo absoluto.

43
Neutralismo Taxa de substituição sob deriva: k = 2Nμ * 1/2N = μ E sob seleção: k = 2N μ * 2s = 4N μ s

44
Neutralismo Previsões da hipótese neutralista: 1- Relógio molecular proporcional ao tempo absoluto? (geracional) (pois proporcional à taxa de mutação). 2- Heterozigose alta, independente do tamanho populacional. 3- Divergência entre populações similar ao polimorfismo dentro das populações.
 (pois proporcional à taxa de mutação). 2- Heterozigose alta, independente do tamanho populacional. 3- Divergência entre populações similar ao polimorfismo dentro das populações..")
45
Heterezigose A taxa de heterozigose tipicamente é ao redor de 0.1 Se H=0.1, como H= 4Nµ / (4Nµ+1) 4Nµ ~ 0.1 Usando µ=5x10 -8 Podemos nos perguntar: qual N necessário? O valor obtido é 500,000 que é razoável.

46
Heterozigose

47
Substituição/polimorfismo pN/pS Sob neutralidade: kN/kS = pN/pS kN/kS pN/pS

48
Substituição/polimorfismo pN/pS Sob seleção positiva kN/kS > pN/pS (Drosophila) kN/kS pN/pS = subst. não sinônima

49
Substituição/polimorfismo pN/pS kN/kS Sob modelo com mutações fracamente deletérias kN/kS < pN/pS (Humanos) pN/pS = polim. não sinônimo

50
Exemplo de baixo coeficiente de seleção

51
Hipótese quase-neutralista Tomoko Ohta “A teoria quase neutra pode ser resumida da seguinte forma. Tanto a deriva genética como a seleção influenciam o comportamento de mutações fracamente selecionadas. A deriva predomina em populações pequenas, e a seleção em populações grandes. A maioria das novas mutações é deletéria, e a maioria das mutações de efeito pequeno devem ser muito fracamente deletérias. Há seleção contra essas mutações em populações grandes, mas se comportam como neutras e populações pequenas”

52
Heterozigose

53
Estimativas de divergência A vida seria fácil com o relógio molecular...

54
Estimativas de divergência

Apresentações semelhantes