Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Bioquímica de frutos Alexandra Mara G. N. Mamede

2
Aula 1 Introdução à Frutas & Hortaliças

3
Fisiologia pós-colheita de frutas e hortaliças
Função específica da planta Estágio de desenvolvimento e maturação Fatores climáticos Práticas agrícolas Manipulação pós-colheita e estocagem Fatores endógenos Fatores exógenos Manipulação pós-colheita: importante a retirada do calor do campo para uma maior vida útil dos produtos hortícolas

4
Frutas e hortaliças Partes comestíveis frutos, folhas, caules, raízes,
Partes da planta Exemplos Raiz Cenoura, nabo, batata doce, mandioca Caule Aspargos Tubérculo Batata, inhame Folha Alface, espinafre, repolho Parte Floral Alcachofra, brócolis, couve-flor Bulbo Cebola, alho Frutos Imaturos não carnosos Ervilha fresca, feijão verde, quiabo, milho verde Maturos não carnosos Sementes e nozes Imaturos carnosos Pepino, abobrinha Maturos carnosos Maçã, pêra, pêssego, uva, citros, melão, tomate, abóbora Partes comestíveis frutos, folhas, caules, raízes, flores, etc. Stem - caule, haste Um bolbo ou bulbo é um tipo de órgão vegetal de algumas plantas perenes que inclui uma parte correspondente ao caule, geralmente de forma discoidal, da qual partem raízes e folhas modificadas escamiformes que servem como órgão de armazenamento de nutrientes que servirão a planta durante a época desfavorável, em que perdem a parte aérea, perdendo, portanto, a capacidade de realizar a fotossíntese. As plantas com bolbos verdadeiros são sempre monocotiledóneas e incluem, por exemplo: A cebola, o alho e todas as Alliaceae. Lírios, tulipas e outras Liliaceae. Amaryllis, Hippeastrum, Narciso, e outras Amaryllidaceae. Dois grupos de 'Iris: Xiphium e Hermodactyloides. Fennema, 2008

5
DEFINIÇÃO DOS FRUTOS “Produtos comestíveis de árvores ou plantas, constituídos de semente(s) e seu invólucro, é geralmente suculento e polpudo” (Dicionário Oxford); O fruto comestível carnoso e adocicado é designado como “fruta”; “Resultado do desenvolvimento do ovário das flores ou inflorescências das angiospermas, em consequência da fecundação do(s) óvulos(s)” (Botanicamente). Pericarpo: Provém das paredes do ovário e consta, geralmente, de três partes: epicarpo, a parte externa e comumente delgada; mesocarpo, a parte mediana por vezes muito espessas; endocarpo, situado internamente e em contato com a semente ou sementes. Angiospermas plantas mais evoluidas que possuem semente e flores Chitarra & Chitarra, 2005
 e seu invólucro, é geralmente suculento e polpudo (Dicionário Oxford); O fruto comestível carnoso e adocicado é designado como fruta ; Resultado do desenvolvimento do ovário das flores ou inflorescências das angiospermas, em consequência da fecundação do(s) óvulos(s) (Botanicamente). Pericarpo: Provém das paredes do ovário e consta, geralmente, de três partes: epicarpo, a parte externa e comumente delgada; mesocarpo, a parte mediana por vezes muito espessas; endocarpo, situado internamente e em contato com a semente ou sementes. Angiospermas plantas mais evoluidas que possuem semente e flores. Chitarra & Chitarra,")
6
CLASSIFICAÇÃO DOS FRUTOS
(Chitarra & Chitarra, 2005) epicarpo Pericarpo: Parte do fruto que envolve a semente. Divide-se em epicarpo (parte mais externa ou casca), mesocarpo (marte intermediária) e endocarpo (parte mais interna que normalmente reveste a semente). Em botânica, chama-se pericarpo à camada externa do fruto das angiospermas, que envolve as sementes. O pericarpo é originado pelo desenvolvimento do ovário, que segue à fecundação. Pode dividir-se em exocarpo ou epicarpo (vulgarmente, casca ou pele) normalmente uma camada membranácea e fibrosa, que pode ser lisa, rugosa, pilosa ou espinosa que protege o mesocarpo, podendo ainda distinguir-se um endocarpo, que igualmente pode ser membranoso ou rígido[1]. Deiscendentes: qdo o pericarpo se abre para liberar as sementes (legumes) Indeiscentes: qdo não se abrem (nozes) Esquizocárpicos: qdo não abrem, porém, as sementes não ficam expostas, como as sâmaras duplas Simples : quando se originam de uma flor de carpelos unidos entre si. Exemplo: Laranja, Maçã, Pera, Tomate,… Compostos : se originam de uma flor com carpelos separados. Exemplo: Morango, Jaca, Fruta do Conde ou Pinha,… Carpelo : é a folha modificada que se fecha sobre os ovários, como se fossem “casinhas em volta dos ovários”. O caju é considerado um Pseudofruto, pois a sua parte suculenta - a parte que comemos- é o pendúculo da Flor e não o ovário desenvolvido. Folha carpelar ou carpelo – a folha carpelar toma a forma de uma garrafa, na qual se podem reconhecer três partes: estigma, estilete e ovário. No interior do ovário formam-se os óvulos.
 epicarpo. Pericarpo: Parte do fruto que envolve a semente. Divide-se em epicarpo (parte mais externa ou casca), mesocarpo (marte intermediária) e endocarpo (parte mais interna que normalmente reveste a semente). Em botânica, chama-se pericarpo à camada externa do fruto das angiospermas, que envolve as sementes. O pericarpo é originado pelo desenvolvimento do ovário, que segue à fecundação. Pode dividir-se em exocarpo ou epicarpo (vulgarmente, casca ou pele) normalmente uma camada membranácea e fibrosa, que pode ser lisa, rugosa, pilosa ou espinosa que protege o mesocarpo, podendo ainda distinguir-se um endocarpo, que igualmente pode ser membranoso ou rígido[1]. Deiscendentes: qdo o pericarpo se abre para liberar as sementes (legumes) Indeiscentes: qdo não se abrem (nozes) Esquizocárpicos: qdo não abrem, porém, as sementes não ficam expostas, como as sâmaras duplas. Simples : quando se originam de uma flor de carpelos unidos entre si. Exemplo: Laranja, Maçã, Pera, Tomate,… Compostos : se originam de uma flor com carpelos separados. Exemplo: Morango, Jaca, Fruta do Conde ou Pinha,… Carpelo : é a folha modificada que se fecha sobre os ovários, como se fossem casinhas em volta dos ovários . O caju é considerado um Pseudofruto, pois a sua parte suculenta - a parte que comemos- é o pendúculo da Flor e não o ovário desenvolvido. Folha carpelar ou carpelo – a folha carpelar toma a forma de uma garrafa, na qual se podem reconhecer três partes: estigma, estilete e ovário. No interior do ovário formam-se os óvulos.")
8
DEFINIÇÃO DAS HORTALIÇAS
Partes de plantas que não pertencem ao grupo de frutas e cereais e que são consumidas frescas, cruas ou processadas (Chitarra & Chitarra, 2005)
")
9
CLASSIFICAÇÃO DAS HORTALIÇAS
Por categoria de órgãos ou partes da planta: Sementes e vagens Bulbos, raízes, rizomas e tubérculos Flores, brotos, hastes e folhas Frutos (pepino, abobrinha, melão) Pelas características morfológicas na planta: Partes aéreas: Folhas (ex. espinafre, alface, couve, repolho, salsa, endívia) Pecíolos (ex. aipo, erva-doce, ruibarbo) Hastes (ex. aspargos, aipo) Inflorescências (ex. brócolis, alcachofra, couve-flor) Deiscendentes: qdo o pericarpo se abre para liberar aas sementes (legumes) Indeiscentes: qdo não se abrem (nozes) Esquizocárpicos: qdo não abrem, porém, as sementes não ficam expostas, como as sâmaras duplas??????
 Pelas características morfológicas na planta: Partes aéreas: Folhas (ex. espinafre, alface, couve, repolho, salsa, endívia) Pecíolos (ex. aipo, erva-doce, ruibarbo) Hastes (ex. aspargos, aipo) Inflorescências (ex. brócolis, alcachofra, couve-flor) Deiscendentes: qdo o pericarpo se abre para liberar aas sementes (legumes) Indeiscentes: qdo não se abrem (nozes) Esquizocárpicos: qdo não abrem, porém, as sementes não ficam expostas, como as sâmaras duplas")
10
CLASSIFICAÇÃO DAS HORTALIÇAS
Pelas características morfológicas na planta: Partes subterrâneas: Raízes (ex. cenoura, beterraba, aipo, nabo, rabanete, batata-doce, mandioca) Rizomas e tubérculos (ex. batata, inhame, gengibre) Bulbos (ex. alho, cebola) Frutos: Imaturos carnosos (ex. abobrinha, berinjela, pepino, pimentão, jiló) Imaturos não carnosos e sementes (ex. ervilha, milho-doce,quiabo, vagem) Frutos maturos: polpudos e macios(ex. tomate e melão) polpudos e duros (ex. abóbora) Sementes (ex. feijões e lentilhas)
 Rizomas e tubérculos (ex. batata, inhame, gengibre) Bulbos (ex. alho, cebola) Frutos: Imaturos carnosos (ex. abobrinha, berinjela, pepino, pimentão, jiló) Imaturos não carnosos e sementes (ex. ervilha, milho-doce,quiabo, vagem) Frutos maturos: polpudos e macios(ex. tomate e melão) polpudos e duros (ex. abóbora) Sementes (ex. feijões e lentilhas)")
11
Tecidos e células vegetais
TIPOS DE TECIDOS DE PROTEÇÃO OU DÉRMICO: casca, epicarpo ou epiderme Proteção contra agressão física, química ou biológica Responsáveis pelas trocas hídricas e gasosas com o meio externo Ricos em material lipídico depositados em camadas (cutícula: ceras, cutina, suberina) FUNDAMENTAL: Parênquima Armazenamento de nutrientes Tecido predominante nas partes macias do vegetal Tecido mais abundante nas plantas comestíveis Ricos em pectinas Recobrem a superficie (tecido dérmico)
 FUNDAMENTAL: Parênquima. Armazenamento de nutrientes. Tecido predominante nas partes macias do vegetal. Tecido mais abundante nas plantas comestíveis. Ricos em pectinas. Recobrem a superficie (tecido dérmico)")
12
DE SUPORTE: Colênquima e esclerênquima
TIPOS DE TECIDOS DE SUPORTE: Colênquima e esclerênquima Proporcionam firmeza com flexibilidade ou dureza ao vegetal Ricos em celulose, hemiceluloses, pectinas e lignina VASCULAR: Xilema e floema Constituído por canais para transporte de água e nutrientes Tecido complexo: algumas células altamente especializadas MERISTEMÁTICO : Células com capacidade de divisão celular

13
Tipos de tecidos (Taiz e Zeiger, 2004)
")
14
Célula As células são as microunidades que formam os organismos vivos, sendo constituídas de uma massa de protoplasma contendo organelas responsáveis pelas transformações metabólicas e reprodução das espécies vegetais.

15
Célula ORGANELAS: (núcleo, vacúolos, mitocôndrias, cloroplastos, etc. ) Especializadas para armazenamento: Polissacarídeos (amiloplastos para amido); Proteína (“corpos proteicos” no endosperma e camada de aleurona); Lipídeos (esferosomas, gotículas de triacilglicerois no endosperma e camada de aleurona de sementes)
; Proteína ( corpos proteicos no endosperma e camada de aleurona); Lipídeos (esferosomas, gotículas de triacilglicerois no endosperma e camada de aleurona de sementes)")
16
Célula PAREDE CELULAR: responsável pela resistência e rigidez dos tecidos vegetais. PRIMÁRIA, SECUNDÁRIA E LAMELA MÉDIA Parede celular primária: As microfibrilas de celulose são revestidas com hemicelulose (como o xiloglucano), que podem também ligar as microfibrilas entre si. As pectinas formam um gel de matriz de ligação, talvez interagindo com proteínas estruturais Hemiceluloses e celulose representam mais de 50% do peso seco de resíduos agrícolas (7), eles podem ser convertidos em açúcares solúveis, quer por hidrólise ácida ou enzimática, de modo a que possam ser utilizados como uma abundante e barata fonte de energia renovável no mundo. (Taiz e Zeiger, 2004)
, que podem também ligar as microfibrilas entre si. As pectinas formam um gel de matriz de ligação, talvez interagindo com proteínas estruturais. Hemiceluloses e celulose representam mais de 50% do peso seco de resíduos agrícolas (7), eles podem ser convertidos em açúcares solúveis, quer por hidrólise ácida ou enzimática, de modo a que possam ser utilizados como uma abundante e barata fonte de energia renovável no mundo. (Taiz e Zeiger, 2004)")
17
(Chitarra & Chitarra, 200)
")
19
Fases do desenvolvimento de frutos
Série de eventos desde o início do crescimento de um fruto até a morte do mesmo.

20
Fases do desenvolvimento de frutos
Pré-maturação: Geralmente inclui a metade do período entre a floração e a colheita. Esse estádio e caracterizado pelo extensivo aumento do volume e termina quando o desenvolvimento do fruto e apenas aceitável, mas não ótimo para o consumo. Maturação Aumento do tamanho até o término do crescimento Sequência de mudanças bioquímicas, fisiológicas e estruturais dos frutos, conduzindo a um estado que os torna comestíveis.

21
Fases do desenvolvimento de frutos
Amadurecimento Torna os frutos em produtos atrativos e aptos para o consumo humano Etapa intermediária entre o final do desenvolvimento e o início da senescência, sendo um processo normal e irreversível Senescência Ocorrem após a maturidade fisiológica ou horticultural Período de predominância dos processos degradativos, que resultam na morte dos tecidos, tornando o fruto inadequado para o consumo Maturidade comercial: indica o ponto de colheita

22
Fases do desenvolvimento de frutos
SUB FASES DA MATURAÇÃO Pré-climatério: etapa da maturação que antecede a elevação súbita da produção de etileno e da atividade respiratória em alguns tipos de frutos. Climatério: corresponde a elevação súbita da produção autocatalítica de etileno e da respiração em alguns tipos de frutas, induzindo ao rápido amadurecimento dos mesmos (frutos climatérios). Pós-climatério: fase de declínio na produção súbita de etileno e na atividade respiratória de alguns tipos de frutos, indicativa do início da senescência.
. Pós-climatério: fase de declínio na produção súbita de etileno e na atividade respiratória de alguns tipos de frutos, indicativa do início da senescência.")
23
Fases do desenvolvimento de frutos

24
Produtos perecíveis Elevada atividade metabólica após a colheita Manutenção da qualidade Conhecimento da estrutura, da fisiologia e das transformações metabólicas no ciclo vital QUALIDADE Conjunto de atributos que determina o grau de aceitação pelo consumidor

25
Características de Qualidade
Produtor Boa produtividade Resistência a pragas e doenças Fácil manejo e fácil colheita Aparência Atacadista e Varejista Firmeza Resistência ao transporte Durabilidade Consumidor Aparência (Cor) Firmeza (Textura) Sabor e Aroma Nutritivo Sem resíduos Fonte: CQH - CEAGESP Características de Qualidade (Conjunto de atributos que determina o grau de aceitação pelo consumidor) Nutritivo entra carboidratos (açúcares, proteínas, ácidos orgânicos, vitaminas, compostos fenólicos)
 Firmeza (Textura) Sabor e Aroma. Nutritivo. Sem resíduos. Fonte: CQH - CEAGESP. Características de Qualidade (Conjunto de atributos que determina o grau de aceitação pelo consumidor) Nutritivo entra carboidratos (açúcares, proteínas, ácidos orgânicos, vitaminas, compostos fenólicos)")
26
TRANSFORMAÇÕES BIOQUÍMICAS APÓS A COLHEITA
Durante o crescimento e a maturação as frutas e hortaliças são dependentes da fotossíntese e da absorção de água e minerais pela planta. Depois de colhidas elas tornam-se unidades independentes e a respiração passa a desempenhar um importante papel. Características de Qualidade (Conjunto de atributos que determina o grau de aceitação pelo consumidor)
")
27
As principais transformações (físicas, químicas e bioquímicas) que ocorrem durante a maturação e refletem nos atributos de qualidade dos produtos hortícolas estão agrupados a seguir: Desenvolvimento das sementes Síntese protéica (enzimas) Modificação na permeabilidade das membranas celulares Elevação da atividade respiratória Síntese de etileno Modificação na pigmentação: degradação da clorofila, com aparecimento de pigmentos pré-existentes; síntese de carotenóides e de flavonóides.
 Modificação na permeabilidade das membranas celulares. Elevação da atividade respiratória. Síntese de etileno. Modificação na pigmentação: degradação da clorofila, com aparecimento de pigmentos pré-existentes; síntese de carotenóides e de flavonóides.")
28
Modificação da textura:
solubilização das pectinas; hidrólise de polissacarídeos estruturais da parede celular. Modificação do sabor e do aroma: hidrólise de polissacarídeos de reserva; interconversão de açúcares; síntese e/ou degradação de ácidos orgânicos; polimerização de fenólicos; síntese de compostos voláteis (aromáticos)
")
29
Aula 2 Atividade Respiratória

30
RESPIRAÇÃO Reações oxidativas de compostos orgânicos que são transformados em água e CO2 com produção de energia química, utilizada para a biossíntese de novos compostos indispensáveis ao perfeito funcionamento e manutenção da planta como um todo. A respiração é um processo primário e primordial de todo ser vivo. No produto vegetal colhido, esse processo também é continuo e se traduz pela quebra oxidativa de materiais orgânicos armazenados (carboidratos, proteínas e gorduras) que são transformados em produtos finais simples, incluindo CO2 e H2O, com produção de energia química, utilizada para a biossíntese de novos compostos indispensáveis ao perfeito funcionamento e manutenção da planta como um todo (Figura 1) (CHITARRA & PRADO, 2002) A energia química liberada através da respiração é captada para dar continuidade aos processos de síntese necessários à sua sobrevivência, notadamente no que se refere à organização celular, à permeabilidade das membranas e ao transporte de metabólicos para os tecidos. Dessa forma, as células continuam a produção de enzimas e de outras substâncias de estrutura molecular elaborada, como parte essencial do processo de manutenção de suas funções vitais. Essas sínteses são parte essencial do amadurecimento de muitos frutos Após a colheita de qualquer parte do vegetal, a respiração torna-se o seu principal processo fisiológico, uma vez que ele não depende mais da absorção de água e minerais efetuados pelas raízes, da condução de nutrientes pelo sistema vascular, nem da atividade fotossintética das folhas da planta mãe. Portanto, após a colheita, as partes do vegetal adquirem vida independente e utilizam, para tal, suas próprias reservas metabólicas acumuladas nas fases de crescimento e de maturação, com conseqüente depressão progressiva nas reservas de matéria seca acumulada A perda de reservas armazenadas no produto durante a respiração é traduzida pelo avanço da senescência à medida que as reservas que proporcionam energia para manter o status de vida do produto são exauridas pela redução do valor alimentar (valor energético) para o consumidor, pela perda da qualidade do sabor e aroma, especialmente doçura e pela perda de peso seco comercializável (KADER, 2002). Portanto, o controle da respiração é condição essencial para a manutenção da qualidade e para o prolongamento da vida útil dos produtos vegetais perecíveis (CHITARRA, 1998).
 que são transformados em produtos finais simples, incluindo CO2 e H2O, com produção de energia química, utilizada para a biossíntese de novos compostos indispensáveis ao perfeito funcionamento e manutenção da planta como um todo (Figura 1) (CHITARRA & PRADO, 2002) A energia química liberada através da respiração é captada para dar continuidade aos processos de síntese necessários à sua sobrevivência, notadamente no que se refere à organização celular, à permeabilidade das membranas e ao transporte de metabólicos para os tecidos. Dessa forma, as células continuam a produção de enzimas e de outras substâncias de estrutura molecular elaborada, como parte essencial do processo de manutenção de suas funções vitais. Essas sínteses são parte essencial do amadurecimento de muitos frutos. Após a colheita de qualquer parte do vegetal, a respiração torna-se o seu principal processo fisiológico, uma vez que ele não depende mais da absorção de água e minerais efetuados pelas raízes, da condução de nutrientes pelo sistema vascular, nem da atividade fotossintética das folhas da planta mãe. Portanto, após a colheita, as partes do vegetal adquirem vida independente e utilizam, para tal, suas próprias reservas metabólicas acumuladas nas fases de crescimento e de maturação, com conseqüente depressão progressiva nas reservas de matéria seca acumulada. A perda de reservas armazenadas no produto durante a respiração é traduzida pelo avanço da senescência à medida que as reservas que proporcionam energia para manter o status de vida do produto são exauridas pela redução do valor alimentar (valor energético) para o consumidor, pela perda da qualidade do sabor e aroma, especialmente doçura e pela perda de peso seco comercializável (KADER, 2002). Portanto, o controle da respiração é condição essencial para a manutenção da qualidade e para o prolongamento da vida útil dos produtos vegetais perecíveis (CHITARRA, 1998).")
31
PRINCIPAL PROCESSO FISIOLÓGICO
APÓS A COLHEITA RESPIRAÇÃO PRINCIPAL PROCESSO FISIOLÓGICO Principal fenômeno que influencia a conservação e qualidade das frutas e hortaliças. A velocidade de respiração é um bom índice para predizer o tempo de vida útil dos produtos hortícolas após a colheita. A respiração e o principal processo fisiológico que continua ocorrendo após a colheita. Nesta etapa, a respiração se realiza graças as reservas acumuladas pela fruta, uma vez que ela não depende mais da absorção de água e nutrientes pelas raízes, e da atividade fotossintética das folhas da planta que a produziu. A perda de reservas armazenadas no produto durante a respiração é traduzida pelo avanço da senescência à medida que as reservas que proporcionam energia para manter o status de vida do produto são exauridas pela redução do valor alimentar (valor energético) para o consumidor, pela perda da qualidade do sabor e aroma, especialmente doçura e pela perda de peso seco comercializável (KADER, 2002). Portanto, o controle da respiração é condição essencial para a manutenção da qualidade e para o prolongamento da vida útil dos produtos vegetais perecíveis (CHITARRA, 1998).
 para o consumidor, pela perda da qualidade do sabor e aroma, especialmente doçura e pela perda de peso seco comercializável (KADER, 2002). Portanto, o controle da respiração é condição essencial para a manutenção da qualidade e para o prolongamento da vida útil dos produtos vegetais perecíveis (CHITARRA, 1998).")
32
RESERVAS ACUMULADAS SÃO SUBSTRATOS
APÓS A COLHEITA RESPIRAÇÃO RESERVAS ACUMULADAS SÃO SUBSTRATOS Não depende mais da absorção de água e nutrientes pelas raízes, e da atividade fotossintética das folhas da planta mãe A respiração e o principal processo fisiológico que continua ocorrendo após a colheita. Nesta etapa, a respiração se realiza graças as reservas acumuladas pela fruta, uma vez que ela não depende mais da absorção de água e nutrientes pelas raízes, e da atividade fotossintética das folhas da planta que a produziu. A perda de reservas armazenadas no produto durante a respiração é traduzida pelo avanço da senescência à medida que as reservas que proporcionam energia para manter o status de vida do produto são exauridas pela redução do valor alimentar (valor energético) para o consumidor, pela perda da qualidade do sabor e aroma, especialmente doçura e pela perda de peso seco comercializável (KADER, 2002). Portanto, o controle da respiração é condição essencial para a manutenção da qualidade e para o prolongamento da vida útil dos produtos vegetais perecíveis (CHITARRA, 1998).
 para o consumidor, pela perda da qualidade do sabor e aroma, especialmente doçura e pela perda de peso seco comercializável (KADER, 2002). Portanto, o controle da respiração é condição essencial para a manutenção da qualidade e para o prolongamento da vida útil dos produtos vegetais perecíveis (CHITARRA, 1998).")
33
RESPIRAÇÃO Principal fenômeno que influencia a conservação e qualidade das frutas e hortaliças. A velocidade de respiração é um bom índice para predizer o tempo de vida útil dos produtos hortícolas após a colheita. A perda de reservas armazenadas no produto durante a respiração é traduzida pelo avanço da senescência à medida que as reservas que proporcionam energia para manter o status de vida do produto são exauridas pela redução do valor alimentar (valor energético) para o consumidor, pela perda da qualidade do sabor e aroma, especialmente doçura e pela perda de peso seco comercializável (KADER, 2002). Portanto, o controle da respiração é condição essencial para a manutenção da qualidade e para o prolongamento da vida útil dos produtos vegetais perecíveis (CHITARRA, 1998).
 para o consumidor, pela perda da qualidade do sabor e aroma, especialmente doçura e pela perda de peso seco comercializável (KADER, 2002). Portanto, o controle da respiração é condição essencial para a manutenção da qualidade e para o prolongamento da vida útil dos produtos vegetais perecíveis (CHITARRA, 1998).")
34
Atividade respiratória e vida de prateleira
nabo ervilhas ervilhas Produtos com taxas de respiração relativamente baixas são os que podem ser armazenados por períodos mais longos sem perda da aceitabilidade (página 81 livro Chitarra) The importance of respiration in postharvest crops is best illustrated by comparing the relationship between respiratory rate and storage life of different crops (Fig. 20). Commodities that exhibit rapid rates of carbon dioxide evolution or oxygen consumption are generally quite perishable, whereas those with slow respiratory rates may be stored satisfactorily for relativelylong periods of time. Moreover, the shelf life of a given commodity can be greatly extended by placing it in an environment that appropriately retards respiration (e.g., refrigeration, or controlled atmosphere). Fennema, 1997
 The importance of respiration in postharvest crops is best illustrated by comparing the relationship between respiratory rate and. storage life of different crops (Fig. 20). Commodities that exhibit rapid rates of carbon dioxide evolution or oxygen consumption. are generally quite perishable, whereas those with slow respiratory rates may be stored satisfactorily for relativelylong periods of time. Moreover, the shelf life of a given commodity can be greatly extended by placing it in an environment that appropriately retards respiration (e.g., refrigeration, or controlled atmosphere). Fennema,")
35
VIAS METABÓLICAS - RESPIRAÇÃO
Glicólise (Embden-Meyerhoff-Parnas) + Ciclo de Krebs (TCA cycle) Produção de ATP Produção de ácidos orgânicos (cítrico, málico) Ciclo das pentoses 6Gli-6P + 12 NADP+ 5 Gli-6P + 6 CO NADPH Produção de NADPH (Processos de Biossíntese: pigmentos e componentes aromáticos) Participação variável das vias A respiração é um processo primário e primordial de todo ser vivo. No produto vegetal colhido, esse processo também é continuo e se traduz pela quebra oxidativa de materiais orgânicos armazenados (carboidratos, proteínas e gorduras) que são transformados em produtos finais simples, incluindo CO2 e H2O, com produção de energia química, utilizada para a biossíntese de novos compostos indispensáveis ao perfeito funcionamento e manutenção da planta como um todo (Figura 1) A energia química liberada é armazenada na forma de ATP (trifosfato de adenosina) que atua como intermediário nos processos metabólicos que requerem energia. A molécula de ATP consiste de um núcleo adenosina (adenina + ribose) ligado a três radicais fosfato (Figura 2) (PAIVA, 2000). Após a colheita, a respiração nos vegetais pode envolver diferentes vias bioquímicas, mas o processo de respiração em si resume-se a uma combustão indireta do carbono na presença de oxigênio. A oxidação completa de l mol de glicose (fonte de carbono) ocorre de acordo com a seguinte equação geral (CHITARRA & CHITARRA, 2005): A respiração é um processo primário e primordial de todo ser vivo. No produto vegetal colhido, esse processo também é continuo e se traduz pela quebra oxidativa de materiais orgânicos armazenados (carboidratos, proteínas e gorduras) que são transformados em produtos finais simples, incluindo CO2 e H2O, com produção de energia química, utilizada para a biossíntese de novos compostos indispensáveis ao perfeito funcionamento e manutenção da planta como um todo (Figura 1) (CHITARRA & PRADO, 2002), (KADER, 2002) e (CHITARRA & CHITARRA, 2005).
 + Ciclo de Krebs (TCA cycle) Produção de ATP. Produção de ácidos orgânicos (cítrico, málico) Ciclo das pentoses. 6Gli-6P + 12 NADP+ 5 Gli-6P + 6 CO NADPH. Produção de NADPH (Processos de Biossíntese: pigmentos e componentes aromáticos) Participação variável das vias. A respiração é um processo primário e primordial de todo ser vivo. No produto vegetal colhido, esse processo também é continuo e se traduz pela quebra oxidativa de materiais orgânicos armazenados (carboidratos, proteínas e gorduras) que são transformados em produtos finais simples, incluindo CO2 e H2O, com produção de energia química, utilizada para a biossíntese de novos compostos indispensáveis ao perfeito funcionamento e manutenção da planta como um todo (Figura 1) A energia química liberada é armazenada na forma de ATP (trifosfato de adenosina) que atua como intermediário nos processos metabólicos que requerem energia. A molécula de ATP consiste de um núcleo adenosina (adenina + ribose) ligado a três radicais fosfato (Figura 2) (PAIVA, 2000). Após a colheita, a respiração nos vegetais pode envolver diferentes vias bioquímicas, mas o processo de respiração em si resume-se a uma combustão indireta do carbono na presença de oxigênio. A oxidação completa de l mol de glicose (fonte de carbono) ocorre de acordo com a seguinte equação geral (CHITARRA & CHITARRA, 2005): A respiração é um processo primário e primordial de todo ser vivo. No produto vegetal colhido, esse processo também é continuo e se traduz pela quebra oxidativa de materiais orgânicos armazenados (carboidratos, proteínas e gorduras) que são transformados em produtos finais simples, incluindo CO2 e H2O, com produção de energia química, utilizada para a biossíntese de novos compostos indispensáveis ao perfeito funcionamento e manutenção da planta como um todo (Figura 1) (CHITARRA & PRADO, 2002), (KADER, 2002) e (CHITARRA & CHITARRA, 2005).")
36
O amido é o principal carboidrato de reserva nos órgãos vegetais e sua hidrólise produz glicose, que, por sua vez, é oxidada nas reações subseqüentes. A interconverção entre amido e sacarose também ocorre em alguns produtos vegetais. A sacarose é o principal açúcar de translocação das folhas para os frutos e é transformada, quando necessário, em glicose e frutose. Pelo processo de isomerização, a frutose transforma-se em glicose, e vice-versa. Na presença do oxigênio, o ácido pirúvico formado a partir da glicose é convertido a outros ácidos orgânicos, dióxido de carbono (CO2) e água (H2O), com liberação de energia química (ATP). Essa é a via principal utilizada pelas frutas e hortaliças para a produção de energia e dos compostos intermediários que são utilizados como fonte de carbono para a síntese de novos compostos, como pigmentos, aminoácidos, ácido ascórbico, fenólicos, compostos voláteis, etc (CHITARRA & CHITARRA, 2005). As proteínas e os lipídeos também podem servir como substratos, porque existe uma interelação nos seus processos metabólicos. Os glicídeos podem ser convertidos em lipídeos e aminoácidos. Os lipídeos podem ser transformados em aminoácidos, porém, dificilmente transformam-se em glicídeos (CHITARRA & CHITARRA, 2005). Esquema simplificado da respiração. C.K. = Ciclo de Krebs; C.T.E. = cadeia de transporte de elétrons (KLUGE et al., 2002).
. Esquema simplificado da respiração. C.K. = Ciclo de Krebs; C.T.E. = cadeia de transporte de elétrons (KLUGE et al., 2002).")
37
Sabores e odores desagradáveis
Morte celular e perda do produto A respiração pode ocorrer na presença de O2 (aeróbica) ou na sua ausência (anaeróbica ou fermentação). Nesse caso, a liberação de energia será reduzida e a produção de álcoois e aldeídos afetará negativamente a qualidade do fruto. A maior parte da atividade respiratória do fruto está localizada nas mitocôndrias, nos quais se situam os sistemas citocrômicos das células (AWAD, 1993). As plantas superiores são organismos aeróbicos obrigatórios, mas também são hábeis para tolerar ambientes com níveis baixos (hipoxia) ou ausência (anoxia) de oxigênio (PAIVA, 2000). As rotas respiratórias citadas utilizam oxigênio e são as preferenciais em frutos e hortaliças. A atmosfera normal é rica em oxigênio, tanto que a quantidade de oxigênio disponível no tecido è ilimitada. Sob várias condições de armazenamento a quantidade de oxigênio na atmosfera pode ser limitada e insuficiente para manter o metabolismo aeróbico pleno (VILAS BOAS, 1999). Em condições atmosféricas adversas onde à concentração de O2 é baixa ou ausente, a respiração mitocondrial (Ciclo de Krebs e cadeia de transporte de elétrons) é inibida e o metabolismo é substituído pela respiração anaeróbica, por meio do qual o ácido pirúvico é convertido a CO2 e acetaldeído (CH3COH) e este, posteriormente, transforma-se em etanol (CH3CH2OH), conforme a equação abaixo (CHITARRA, 2000) e (PAIVA, 2000). A conseqüência imediata da respiração anaeróbia é o processo denominado fermentação, que provoca modificações no sabor, aroma, cor e textura de frutas e hortaliças vegetal (HONÓRIO & MORETTI, 2002). O acúmulo de acetaldeído e de etanol, em níveis tóxicos, causa à morte celular e a perda do produto. A exposição dos vegetais, a anaerobiose mesmo por curtos períodos, resulta no desenvolvimento de sabores e odores desagradáveis, os quais, podem ou não ser eliminados após o seu retorno à condição de aerobiose. Esquema geral do processo respiratório (aeróbico e anaeróbico) (CHITARRA & CHITARRA, 2005).
 ou na sua ausência (anaeróbica ou fermentação). Nesse caso, a liberação de energia será reduzida e a produção de álcoois e aldeídos afetará negativamente a qualidade do fruto. A maior parte da atividade respiratória do fruto está localizada nas mitocôndrias, nos quais se situam os sistemas citocrômicos das células (AWAD, 1993). As plantas superiores são organismos aeróbicos obrigatórios, mas também são hábeis para tolerar ambientes com níveis baixos (hipoxia) ou ausência (anoxia) de oxigênio (PAIVA, 2000). As rotas respiratórias citadas utilizam oxigênio e são as preferenciais em frutos e hortaliças. A atmosfera normal é rica em oxigênio, tanto que a quantidade de oxigênio disponível no tecido è ilimitada. Sob várias condições de armazenamento a quantidade de oxigênio na atmosfera pode ser limitada e insuficiente para manter o metabolismo aeróbico pleno (VILAS BOAS, 1999). Em condições atmosféricas adversas onde à concentração de O2 é baixa ou ausente, a respiração mitocondrial (Ciclo de Krebs e cadeia de transporte de elétrons) é inibida e o metabolismo é substituído pela respiração anaeróbica, por meio do qual o ácido pirúvico é convertido a CO2 e acetaldeído (CH3COH) e este, posteriormente, transforma-se em etanol (CH3CH2OH), conforme a equação abaixo (CHITARRA, 2000) e (PAIVA, 2000). A conseqüência imediata da respiração anaeróbia é o processo denominado fermentação, que provoca modificações no sabor, aroma, cor e textura de frutas e hortaliças vegetal (HONÓRIO & MORETTI, 2002). O acúmulo de acetaldeído e de etanol, em níveis tóxicos, causa à morte celular e a perda do produto. A exposição dos vegetais, a anaerobiose mesmo por curtos períodos, resulta no desenvolvimento de sabores e odores desagradáveis, os quais, podem ou não ser eliminados após o seu retorno à condição de aerobiose. Esquema geral do processo respiratório (aeróbico e anaeróbico) (CHITARRA & CHITARRA, 2005).")
38
Saldo de 2 moles de ATP e 2 moles de NADH para cada mol de glicose
O piruvato é convertido a acetaldeído e CO2 pela enzima piruvato carboxilase na presença do cofator tiaminapirofosfato (TPP). O acetaldeído é convertido a etanol pela enzima álcool desidrogenase. Na maioria das plantas os principais produtos da fermentação são o CO2 e etanol. Porém algum lactato pode ser formado, principalmente nos estágios mais recentes de anoxia. Contudo a presença de lactato reduz o pH do citoplasma, o que ativa a piruvato descarboxilase e inicia a produção de etanol (PAIVA, 2000). O acúmulo de acetaldeído e de etanol, em níveis tóxicos, causa à morte celular e a perda do produto. A exposição dos vegetais, a anaerobiose mesmo por curtos períodos, resulta no desenvolvimento de sabores e odores desagradáveis, os quais, podem ou não ser eliminados após o seu retorno à condição de aerobiose. Reações da glicólise e fermentação vegetais. As setas duplas indicam reações reversíveis e as simples reações essencialmente irreversíveis (TAIZ et al., 2004). Saldo de 2 moles de ATP e 2 moles de NADH para cada mol de glicose
. O acetaldeído é convertido a etanol pela enzima álcool desidrogenase. Na maioria das plantas os principais produtos da fermentação são o CO2 e etanol. Porém algum lactato pode ser formado, principalmente nos estágios mais recentes de anoxia. Contudo a presença de lactato reduz o pH do citoplasma, o que ativa a piruvato descarboxilase e inicia a produção de etanol (PAIVA, 2000). O acúmulo de acetaldeído e de etanol, em níveis tóxicos, causa à morte celular e a perda do produto. A exposição dos vegetais, a anaerobiose mesmo por curtos períodos, resulta no desenvolvimento de sabores e odores desagradáveis, os quais, podem ou não ser eliminados após o seu retorno à condição de aerobiose. Reações da glicólise e fermentação vegetais. As setas duplas indicam reações reversíveis e as simples reações essencialmente irreversíveis. (TAIZ et al., 2004). Saldo de 2 moles de ATP e 2 moles de NADH para cada mol de glicose.")
39
Esta etapa da respiração tem a finalidade de oxidar completamente o piruvato a CO2 e água.
Ácido Cítrico Ácido Málico Ciclo de Krebs ou Ciclo dos Ácidos Tricarboxílcos (TAIZ et al., 2004).
.")
40
Compostos com anéis aromáticos
Uma via alternativa no metabolismo dos açúcares é a via oxidativa das pentoses-fosfato, também conhecido como via das hexoses-monofosfato (HMP). É uma fonte importante do hidrogênio (poder redutor) e dos intermediários requeridos para a síntese de substâncias formadas durante o amadurecimento e que vão caracterizar o fruto, de acordo com a espécie ou cultivar. Nessa via, pela oxidação de um mol de glicose-6-fosfato a CO2, há produção de 12 moles de NADPH (Figura 9), usado como agente redutor nos processos biossintéticos, quando não ocorre a fotossíntese (diferenciação dos tecidos, germinação de sementes, ausência de luz, entre outros). A importância relativa da rota das pentoses parece aumentar nos tecidos maturos, sendo operativo em frutos como o pimentão, tomate, pepino, laranja e banana. As hexoses-fosfato são provenientes da sacarose e, no final, excedem a concentração dessa. Portanto, a biossíntese da sacarose durante o amadurecimento constitui um importante depósito para a produção de ATP. A via alternativa das pentoses tem como produtos principais, além do NADPH, a ribose-5P, que é utilizada para a biossíntese de ácidos nucléicos, e a eritrose-4P, requerida para a síntese do ácido shiquímico, precursor de compostos com anéis aromáticos (CHITARRA & CHITARRA, 2005). Compostos com anéis aromáticos Via oxidativa das pentoses - fosfato ou das hexoses-monofosfato (HMP) (CHITARRA & CHITARRA, 2005).
. É uma fonte importante do hidrogênio (poder redutor) e dos intermediários requeridos para a síntese de substâncias formadas durante o amadurecimento e que vão caracterizar o fruto, de acordo com a espécie ou cultivar. Nessa via, pela oxidação de um mol de glicose-6-fosfato a CO2, há produção de 12 moles de NADPH (Figura 9), usado como agente redutor nos processos biossintéticos, quando não ocorre a fotossíntese (diferenciação dos tecidos, germinação de sementes, ausência de luz, entre outros). A importância relativa da rota das pentoses parece aumentar nos tecidos maturos, sendo operativo em frutos como o pimentão, tomate, pepino, laranja e banana. As hexoses-fosfato são provenientes da sacarose e, no final, excedem a concentração dessa. Portanto, a biossíntese da sacarose durante o amadurecimento constitui um importante depósito para a produção de ATP. A via alternativa das pentoses tem como produtos principais, além do NADPH, a ribose-5P, que é utilizada para a biossíntese de ácidos nucléicos, e a eritrose-4P, requerida para a síntese do ácido shiquímico, precursor de compostos com anéis aromáticos (CHITARRA & CHITARRA, 2005). Compostos com anéis aromáticos. Via oxidativa das pentoses - fosfato ou das hexoses-monofosfato (HMP) (CHITARRA & CHITARRA, 2005).")
41
A cadeia de transporte de elétrons catalisa um fluxo de elétrons desde o NADH (ou FADH) até o oxigênio, o aceptor final de elétrons do processo respiratório Para cada molécula de glicose oxidada na glicólise e ciclo de Krebs, duas moléculas de NADH são geradas no citoplasma, enquanto que oito moléculas de NADH mais duas moléculas de FADH são produzidas na matriz mitocondrial (PAIVA, 2000). A cadeia de transporte de elétrons (Figura 11) catalisa um fluxo de elétrons desde o NADH (ou FADH) até o oxigênio, o aceptor final de elétrons do processo respiratório (PAIVA, 2000). Fosforilação oxidativa e transporte de elétrons na cadeia respiratória (CHITARRA & CHITARRA, 2005).
. A cadeia de transporte de elétrons (Figura 11) catalisa um fluxo de elétrons desde o NADH (ou FADH) até o oxigênio, o aceptor final de elétrons do processo respiratório (PAIVA, 2000). Fosforilação oxidativa e transporte de elétrons na cadeia respiratória (CHITARRA & CHITARRA, 2005).")
42
Para cada molécula de glicose oxidada na glicólise e ciclo de Krebs, duas moléculas de NADH são geradas no citoplasma, enquanto que oito moléculas de NADH mais duas moléculas de FADH são produzidas na matriz mitocondrial. (CHITARRA & CHITARRA, 2005).
.")
43
Isto é, a respiração celular além do fornecimento de energia (ATP) possui ainda uma outra finalidade que é o fornecimento de esqueletos de carbono ou compostos intermediários para a síntese de outros importantes constituintes celulares. Esses compostos incluem, aminoácidos que podem ser utilizados na síntese de proteínas, nucleotídeos para ácidos nucléicos, e ainda esqueletos de carbono precursores de pigmentos, lipídeos, e alguns compostos aromáticos como as ligninas (Figura 3) (PAIVA, 2000). Após a colheita de qualquer parte do vegetal, a respiração torna-se o seu principal processo fisiológico, uma vez que ele não depende mais da absorção de água e minerais efetuados pelas raízes, da condução de nutrientes pelo sistema vascular, nem da atividade fotossintética das folhas da planta mãe. Portanto, após a colheita, as partes do vegetal adquirem vida independente e utilizam, para tal, suas próprias reservas metabólicas acumuladas nas fases de crescimento e de maturação, com conseqüente depressão progressiva nas reservas de matéria seca acumulada (CHITARRA & CHITARRA, 2005). A energia química liberada através da respiração é captada para dar continuidade aos processos de síntese necessários à sua sobrevivência, notadamente no que se refere à organização celular, à permeabilidade das membranas e ao transporte de metabólicos para os tecidos. Dessa forma, as células continuam a produção de enzimas e de outras substâncias de estrutura molecular elaborada, como parte essencial do processo de manutenção de suas funções vitais. Essas sínteses são parte essencial do amadurecimento de muitos frutos Formação de vários compostos a partir da cadeia respiratória (KLUGE et al., 2002).
. A energia química liberada através da respiração é captada para dar continuidade aos processos de síntese necessários à sua sobrevivência, notadamente no que se refere à organização celular, à permeabilidade das membranas e ao transporte de metabólicos para os tecidos. Dessa forma, as células continuam a produção de enzimas e de outras substâncias de estrutura molecular elaborada, como parte essencial do processo de manutenção de suas funções vitais. Essas sínteses são parte essencial do amadurecimento de muitos frutos. Formação de vários compostos a partir da cadeia respiratória (KLUGE et al., 2002).")
44
Indicadores fisiológicos da atividade respiratória pós-colheita de vegetais
Taxa de consumo de O2 (ex: L/kg.h) Taxa de liberação de CO2 (ex: L/kg.h) Quociente Respiratório: CO2/ O2 Indicadores quantitativos Indicador qualitativo A intensidade ou taxa respiratória é um dos fatores determinantes do potencial de longevidade dos produtos hortícolas durante a pós-colheita. De maneira geral, quanto maior a respiração, menor é o potencial de armazenamento refrigerado e menor a vida útil dos mesmos (KLUGE et al., 2002). Grande parte da energia requerida pelos frutos e hortaliças é suprida pela respiração aeróbica. Esta corresponde ao processo oxidativo por meio do qual as células vivas utilizam suas reservas metabólicas como fonte de carbono para a síntese de novos compostos, com produção de energia. Pode ser medida pela quantidade de oxigênio consumido e/ou de dióxido de carbono liberado pelo produto (mg O2 ou CO2.Kg-1.h-1), conforme apresentado na Figura 4 (CHITARRA & CHITARRA, 2005) e (VILAS BOAS, 1999).
 Taxa de liberação de CO2 (ex: L/kg.h) Quociente Respiratório: CO2/ O2. Indicadores. quantitativos. Indicador. qualitativo. A intensidade ou taxa respiratória é um dos fatores determinantes do potencial de longevidade dos produtos hortícolas durante a pós-colheita. De maneira geral, quanto maior a respiração, menor é o potencial de armazenamento refrigerado e menor a vida útil dos mesmos (KLUGE et al., 2002). Grande parte da energia requerida pelos frutos e hortaliças é suprida pela respiração aeróbica. Esta corresponde ao processo oxidativo por meio do qual as células vivas utilizam suas reservas metabólicas como fonte de carbono para a síntese de novos compostos, com produção de energia. Pode ser medida pela quantidade de oxigênio consumido e/ou de dióxido de carbono liberado pelo produto (mg O2 ou CO2.Kg-1.h-1), conforme apresentado na Figura 4 (CHITARRA & CHITARRA, 2005) e (VILAS BOAS, 1999).")
45
INFLUÊNCIA DO SUBSTRATO NO QUOCIENTE RESPIRATÓRIO
Quociente respiratório – pág 92 Chitarra (CHITARRA & CHITARRA, 2005).
.")
46
RESPIRAÇÃO E INJÚRIA Vitti et al. (2004)
Raízes intactas apresentaram a menor taxa respiratória (5 mL CO2 kg-1 h-1), enquanto beterrabas raladas e descascadas obtiveram taxa respiratória de 30 mL CO2 kg-1 h-1 após quatro horas do processamento. O dano mecânico causado pelo corte ou descascamento é um dos maiores obstáculos na conservação dos produtos minimamente processados, e a taxa respiratória destes produtos é cerca de três a cinco vezes maior que a dos órgãos intactos (Chitarra, 1998). O pico respiratório observado nas beterrabas minimamente processadas é oriundo do estresse provocado pelo corte o qual provoca uma perda da compartimentação celular e, com isto, os substratos do metabolismo respiratório entram em maior contato com os complexos enzimáticos resultando em aumento da taxa respiratória. A redução subseqüente na taxa respiratória das beterrabas minimamente processadas, a partir do quarto dia, é discutível. É possível que haja uma auto-regulação da atividade respiratória do tecido causada pela grande produção de ATP (Purvis, 1997). Outra hipótese é que esta redução da taxa respiratória ocorra pelo fato de os substratos respiratórios pararem de reagir com as enzimas presentes nas células da superfície de corte. Portanto, a taxa respiratória observada a partir do quarto dia pode ser resultante das células localizadas abaixo da superfície danificada pelo corte. Vitti et al. (2004)
, enquanto beterrabas raladas e descascadas obtiveram taxa respiratória de 30 mL CO2 kg-1 h-1 após quatro horas do processamento. O dano mecânico causado pelo corte ou descascamento é um dos maiores obstáculos na conservação dos produtos minimamente processados, e a taxa respiratória destes produtos é cerca de três a cinco vezes maior que a dos órgãos intactos (Chitarra, 1998). O pico respiratório observado nas beterrabas minimamente processadas é oriundo do estresse provocado pelo corte o qual provoca uma perda da compartimentação celular e, com isto, os substratos do metabolismo respiratório entram em maior contato com os complexos enzimáticos resultando em aumento da taxa respiratória. A redução subseqüente na taxa respiratória das beterrabas minimamente processadas, a partir do quarto dia, é discutível. É possível que haja uma auto-regulação da atividade respiratória do tecido causada pela grande produção de ATP (Purvis, 1997). Outra hipótese é que esta redução da taxa respiratória ocorra pelo fato de os substratos respiratórios pararem de reagir com as enzimas presentes nas células da superfície de corte. Portanto, a taxa respiratória observada a partir do quarto dia pode ser resultante das células localizadas abaixo da superfície danificada pelo corte. Vitti et al. (2004)")
47
PADRÕES DE ATIVIDADE RESPIRATÓRIA EM FRUTOS
Climatéricos Ligeiro declínio inicial da atividade respiratória seguida de rápido e acentuado aumento e posterior declínio associado à senescência. Aumento acentuado da síntese de etileno precede ou é simultânea ao pico climatérico. Não Climatéricos Declínio gradual da atividade respiratória sem aumento da síntese de etileno A intensidade ou taxa respiratória é um dos fatores determinantes do potencial de longevidade dos produtos hortícolas durante a pós-colheita. De maneira geral, quanto maior a respiração, menor é o potencial de armazenamento refrigerado e menor a vida útil dos mesmos (KLUGE et al., 2002).
.")
48
Padrões de respiração das frutas: (1) não climatérico; (2) climatérico (KLUGE et al., 2002).
 não climatérico; (2) climatérico (KLUGE et al., 2002).")
49
Frutas Não Climatéricas
CLASSIFICAÇÃO DE ALGUMAS FRUTAS DE ACORDO COM O PADRÃO DE ATIVIDADE RESPIRATÓRIA NO AMADURECIMENTO. Nome Comum Nome Científico Frutas Climatéricas Abacate Persea americana, Mill. Ameixa Prunus domestica, L. Banana Musa sp. Caqui Diospyros kaki, L.f. Figo comum Ficus carica, L. Goiaba Psidium guajava, L. Graviola Annona cherimoya, Mill. Maçã Malus sylvestris, Mill. Mamão Carica papaya, L. Manga Mangifera indica, L. Maracujá Passijlora edulis, Sims. Melancia Citrullus lunatus (Thunb) Mansf. Melão Cantaloupe Cucumis melo, L. (Cantalupensis) Pêssego Prunus persica (L.) Batsch Pêra Pirus communis, L. Frutas Não Climatéricas Azeitona Olea europaea, L. Cacau Theobroma cacao, L. Caju Anacardium occidentale, L. Laranja Citrus sinensis (L.) Osbeck Limão Citrus limon (L.) Burm. f. Morango Fragaria x Ananassa, Duchesne Uva Vitis vinifera, L. (Chitarra & Chitarra, 2005)
 Mansf. Melão Cantaloupe. Cucumis melo, L. (Cantalupensis) Pêssego. Prunus persica (L.) Batsch. Pêra. Pirus communis, L. Frutas Não Climatéricas. Azeitona. Olea europaea, L. Cacau. Theobroma cacao, L. Caju. Anacardium occidentale, L. Laranja. Citrus sinensis (L.) Osbeck. Limão. Citrus limon (L.) Burm. f. Morango. Fragaria x Ananassa, Duchesne. Uva. Vitis vinifera, L. (Chitarra & Chitarra, 2005)")
50
Classificação das hortaliças de acordo com a intensidade da atividade respiratória a 10°C.
Classe Respiração (mg CO2.Kg-1.h-1) Produto Muito baixa < 10 alho Baixa 10-20 pepino, melão, repolho, beterraba, tomate Moderada 20-40 cenoura, aipo, pimentão Elevada 40-70 aspargos, chicória, alface Muito elevada 70-100 feijões, cogumelo, espinafre Extremamente elevada > 100 brócolis, ervilha, salsa, milho-doce Fonte: WEISCHMANN, (1987) citado por (CHITARRA & CHITARRA, 2005).
 Produto. Muito baixa. < 10. alho. Baixa pepino, melão, repolho, beterraba, tomate. Moderada cenoura, aipo, pimentão. Elevada aspargos, chicória, alface. Muito elevada feijões, cogumelo, espinafre. Extremamente elevada. > 100. brócolis, ervilha, salsa, milho-doce. Fonte: WEISCHMANN, (1987) citado por (CHITARRA & CHITARRA, 2005).")
51
PADRÃO RESPIRATÓRIO Fennema, 2008

52
ALGUNS FATORES QUE AFETAM A RESPIRAÇÃO
Espécie e Cultivar; Tipo e parte do vegetal Cobertura superficial Estádio de desenvolvimento Produção endógena de etileno Temperatura, Composição atmosférica ([CO2, O2 e etileno]) Umidade relativa Danos físicos Aplicação exógena de etileno fatores intrínsicos do produto fatores extrínsecos (ambiente) A atividade respiratória varia com a espécie, a cultivar, o tipo de tecido e também com os fatores externos, notadamente com a temperatura, concentração de gases na atmosfera (O2, CO2, C2H4), umidade relativa e com a injúria mecânica dos tecidos. Quanto mais rápida e elevada for à atividade respiratória, menor será o período de vida útil do produto vegetal (CHITARRA, 2000).
 Umidade relativa. Danos físicos. Aplicação exógena de etileno. fatores intrínsicos do produto. fatores extrínsecos. (ambiente) A atividade respiratória varia com a espécie, a cultivar, o tipo de tecido e também com os fatores externos, notadamente com a temperatura, concentração de gases na atmosfera (O2, CO2, C2H4), umidade relativa e com a injúria mecânica dos tecidos. Quanto mais rápida e elevada for à atividade respiratória, menor será o período de vida útil do produto vegetal (CHITARRA, 2000).")
54
A respiração é regulada por ação catalítica de enzimas
A respiração é regulada por ação catalítica de enzimas. Essas são sensíveis à temperatura e aumentam suas atividades de 2 a 4 vezes para cada aumento de 10°C de temperatura até à temperatura de 30°C. Acima desta temperatura a taxa respiratória começa a diminuir, acontecendo a morte do produto por altas temperaturas. A temperatura, portanto, passa a ser o fator determinante no armazenamento de produtos vegetais (CHITARRA & PRADO, 2002).
.")
56
Aula 3 Síntese da Sacarose & Amido; Ácidos Orgânicos; Fitormônios

57
Síntese de sacarose e amido

58
Sacarose Amido Glicose -1- Fosfato Maltose (dissacarídeo)
fosforilase α e β amilase invertase sacarose sintase Glicose -1- Fosfato Maltose (dissacarídeo) UDP-glicose + Frutose Glicose -1- Fosfato maltase fosfoglicomutase hexoquinase Glicose Glicose -6- Fosfato Glicose -6- Fosfato Síntese de parede celular hexose isomerase + ADP ATP hexose isomerase hexoquinase Frutose Frutose -6- Fosfato Frutose -6- Fosfato ADP ATP Glicólise Glicólise Degradação do amido e sacarose para dar origem às hexoses glicose e frutose (WILLS et al., 1998).
 UDP-glicose + Frutose. Glicose -1- Fosfato. maltase. fosfoglicomutase. hexoquinase. Glicose. Glicose -6- Fosfato. Glicose -6- Fosfato. Síntese de parede celular. hexose. isomerase. + ADP. ATP. hexose. isomerase. hexoquinase. Frutose. Frutose -6- Fosfato. Frutose -6- Fosfato. ADP. ATP. Glicólise. Glicólise. Degradação do amido e sacarose para dar origem às hexoses glicose e frutose (WILLS et al., 1998).")
59
CLOROPLASTO MALTOSE 3 4 AMIDO GLICOSE 2 1 GLICOSE-1-P 5 GLICOSE-6-P 7 TRIOSE-6-P 6 CITOPLASMA TRIOSE-6-P Degradação do amido (1. Amido fosforilase; 2. α-amilase; 3. β-amilase; 4. α –glucosidase; 5. fosfoglicomutase; 6. transportador de fosfato) (SEYMOUR et al., 1993)
 (SEYMOUR et al., 1993)")
60
Síntese e Metabolismo de Ácidos Orgânicos
Liberação dos vacúolos por aumento da permeabilidade das membranas Metabólitos Intermediários do: Ciclo de Krebs Via do ácido xiquímico formando ácidos orgânicos (ác.quínico, ac. xiquímico, ac. cinámicos) precursores de Aa aromáticos (fenilalanina e tirosina), antocianinas e ligninas Reações de descarboxilação favorescidas
 precursores de Aa aromáticos (fenilalanina e tirosina), antocianinas e ligninas. Reações de descarboxilação favorescidas.")
61
Fennema, 2008

62
Fennema, 2008

63
Via do Ácido Chiquímico Ácído Quínico Ácído Chiquímico Fennema, 2008

64
Mudanças em açúcares e ácidos orgânicos na maturação de frutas
Açúcares: conversão de amido em açúcares favorecida bem como interconversão de açúcares. Síntese e metabolismo (utilização) de ac.orgãnicos se modifica,compadrão típico para cada fruto to Mudanças nos ácidos durante o amadurecimento de pêras Mudanças nos açúcares durante o amadurecimento de pêras Eskin, 1990 64
 de ac.orgãnicos se modifica,compadrão típico para cada fruto. to. Mudanças nos ácidos durante o amadurecimento de pêras. Mudanças nos açúcares durante o amadurecimento de pêras. Eskin,")
65
HORMÔNIOS DE PLANTAS (FITORMÔNIOS)
ETILENO Hormônio vegetal gasoso Estimula a atividade respiratória Síntese é autocatalítica “Dispara” maturação favorecendo o rápido amadurecimento Considerado o hormônio do amadurecimento Acelera o processo de senescência Hormônio vegetal: é um composto orgânico, não nutriente, produzido na planta, o qual em baixas concentrações, promove, inibe ou modifica processos morfológicos e fisiológicos do vegetal. O etileno, composto volátil de dois carbonos, é produzido endogenamente, talvez, por todas as plantas. Em concentrações tão baixas quanto 0,1microL/L, o etileno pode induzir uma série de respostas fisiológicas, incluindo amadurecimento, senescência e desordens fisiológicas (WATADA, 1986). CH2=CH2
. CH2=CH2.")
66
Biossíntese do etileno
Precursor principal: metionina Uso de metionina marcada (14C) comprovou síntese de etileno em maçã (Lieberman et al, 1966) Síntese autocatalítica Produção de etileno inibida com armazenamento em atmosfera de nitrogênio. Exposição ao oxigênio reativa produção de etileno
 comprovou síntese de etileno em maçã (Lieberman et al, 1966) Síntese autocatalítica. Produção de etileno inibida com armazenamento em atmosfera de nitrogênio. Exposição ao oxigênio reativa produção de etileno.")
67
Biossíntese do etileno
CO2 O2 CO2 A redução de teores de oxigênio abaixo de 8% diminui a produção de etileno pelas frutas frescas e vegetais. O oxigênio é necessário para a produção e ação do etileno. Sob condições anaeróbicas, a conversão de 1ácido carboxílico1aminociclopropano (ACC) a etileno é inibida, resultando na acumulação de ACC no tecido, já que os passos iniciais da biossíntese de etileno, a partir da metionina ocorrem na ausência de O2 (WATADA,1986). Em elevadas concentrações de CO2 pode haver aumento na produção de etileno devido a ocorrência de injúrias fisiológicas no tecido.??????????? Todavia, o mecanismo de indução da produção de etileno não é bem conhecido (WATADA, 1986). O CO2 previne ou retarda os efeitos prejudiciais do etileno em frutas e vegetais frescos, tais como a perda de firmeza e a incidência de desordens fisiológicas. O modo de ação do CO2 na inibição ou redução dos efeitos do etileno ainda não está clara, mas BURG & BURG (1967), citados por KADER (1986), sugerem que o CO2 compete com o etileno pelo sítio de ligação. A produção de etileno por frutas e hortaliças processadas pode ser incrementada em até 20 vezes, quando comparada com vegetal intacto (VAROQUAUX e WILEY, 1997). Detoxificação do cianeto pela ciano-alanino sintase à partir de cisteína (TAIZ et al., 2004).
 a etileno é inibida, resultando. na acumulação de ACC no tecido, já que os passos iniciais da biossíntese de etileno, a partir da metionina ocorrem na ausência de O2 (WATADA,1986). Em elevadas concentrações de CO2 pode haver aumento na produção de etileno devido a ocorrência de injúrias fisiológicas no tecido. Todavia, o mecanismo de indução da produção de etileno não é bem conhecido (WATADA, 1986). O CO2 previne ou retarda os efeitos prejudiciais do etileno em frutas e vegetais frescos, tais como a perda de firmeza e a incidência de desordens. fisiológicas. O modo de ação do CO2 na inibição ou redução dos efeitos do etileno ainda não está clara, mas BURG & BURG (1967), citados por. KADER (1986), sugerem que o CO2 compete com o etileno pelo sítio de ligação. A produção de etileno por frutas e hortaliças processadas pode ser incrementada em até 20 vezes, quando comparada com vegetal intacto. (VAROQUAUX e WILEY, 1997). Detoxificação do cianeto. pela ciano-alanino sintase à partir de cisteína. (TAIZ et al., 2004).")
68
O acúmulo de ACC antecede o pico de etileno
(Chitarra & Chitarra, 2005)
")
69
Aumenta expressão gênica de enzimas do amadurecimento:
Efeitos do etileno Aumenta expressão gênica de enzimas do amadurecimento: Clorofilase Celulase Poligalacturonase (PG) Pectinametilesterase (PME) Fenilalanina amônio-liase (FAL) ACC sintase Piruvato desidrogenase HARVEY (1925; 1928), citado por WATADA (1986), estudou o uso de etileno em várias frutas e verificou perda de clorofila, desenvolvimento de cor de maturação, amolecimento dos tecidos, desenvolvimento de sabor e início da atividade da enzima proteoclástica. O etileno estimula a síntese de enzimas envolvidas com a maturação das frutas e pode causar a perda de firmeza das mesmas, provavelmente, devido a ativação de enzimas que hidrolisam a parede celular (WATADA, 1986). Também está relacionado com o incremento da descoloração da cor verde de vegetais folhosos (LOUGHEED, MURR & TOIVONEN, 1987), talvez devido ao aumento da atividade da clorofilase (VAROQUAUX & WILEY, 1997). Além dos metabólitos primários, uma grande variedade de metabólitos secundários é sintetizada ao longo do biociclo vegetal, para os quais muitas funções ecológicas, fisiológicas e curativas têm sido atribuídas (Kouki & Manetas, 2002). Segundo Santos e Palazzo de Melo (2000), os taninos são substâncias polifenólicas de alto peso molecular e sabor adstringente, encontrados no interior dos vacúolos vegetais. Além de sua larga utilização nas indústrias de curtume, tradicionalmente são também empregados na medicina popular, devido a sua ação adstringente (Haslam, 1996). De acordo com De Bruyne et al. (1999), os taninos, assim como os demais compostos fenólicos, derivam-se de um intermediário biossintético comum, a fenilalanina, ou de seu precursor, o ácido chiquímico, sintetizados através da Rota do Chiquimato A fenilalanina amônia-liase destaca-se como uma enzima chave e regulatória da rota de biossíntese dos fenilpropanóides e seus derivados, catalisando a transformação, por desaminação, do aminoácido Lfenilalanina, em ácido trans-cinâmico, sendo este o primeiro passo para a biossíntese dos fenólicos vegetais (Cheng et al., 2001).
 Pectinametilesterase (PME) Fenilalanina amônio-liase (FAL) ACC sintase. Piruvato desidrogenase. HARVEY (1925; 1928), citado por WATADA (1986), estudou o uso de etileno em várias frutas e verificou perda de clorofila, desenvolvimento de cor de maturação, amolecimento dos tecidos, desenvolvimento de sabor e início da atividade da enzima proteoclástica. O etileno estimula a síntese de enzimas envolvidas com a maturação das frutas e pode causar a perda de firmeza das mesmas, provavelmente, devido a ativação de enzimas que hidrolisam a parede celular (WATADA, 1986). Também está relacionado com o incremento da descoloração da cor verde de vegetais folhosos (LOUGHEED, MURR & TOIVONEN, 1987), talvez devido ao aumento da atividade da clorofilase (VAROQUAUX & WILEY, 1997). Além dos metabólitos primários, uma grande. variedade de metabólitos secundários é sintetizada. ao longo do biociclo vegetal, para os quais muitas. funções ecológicas, fisiológicas e curativas têm sido. atribuídas (Kouki & Manetas, 2002). Segundo Santos. e Palazzo de Melo (2000), os taninos são substâncias. polifenólicas de alto peso molecular e sabor. adstringente, encontrados no interior dos vacúolos. vegetais. Além de sua larga utilização nas indústrias. de curtume, tradicionalmente são também. empregados na medicina popular, devido a sua ação. adstringente (Haslam, 1996). De acordo com De. Bruyne et al. (1999), os taninos, assim como os. demais compostos fenólicos, derivam-se de um intermediário biossintético comum, a fenilalanina, ou. de seu precursor, o ácido chiquímico, sintetizados. através da Rota do Chiquimato. A fenilalanina amônia-liase destaca-se como. uma enzima chave e regulatória da rota de biossíntese. dos fenilpropanóides e seus derivados, catalisando a. transformação, por desaminação, do aminoácido Lfenilalanina, em ácido trans-cinâmico, sendo este o. primeiro passo para a biossíntese dos fenólicos. vegetais (Cheng et al., 2001).")
70
EFEITO DO ETILENO EXÓGENO NA ATIVIDADE RESPIRATÓRIA DE FRUTOS CLIMATÉRICOS E NÃO CLIMATÉRICOS
Se aplicar etileno em frutos climatéricos a ação é irreversível, o etileno estimula o climatério respiratório. Quando aplicado no fruto não climatérico o etileno provoca ação reversível Aplicação de etileno em frutos não climatéricos Estimula a respiração em qualquer estádio de maturação Reação reversível Aumenta degradação clorofila (clorofilase) Aumenta síntese de pigmentos (Carotenóides em citros, Antocianina em uva) Aplicação de etileno em frutos climatéricos Estimula a respiração antes da ascensão respiratória, no pré-climatérico Reação irreversível Aumenta enzimas ligadas ao amadurecimento (Kays,1991)
 Aumenta síntese de pigmentos (Carotenóides em citros, Antocianina em uva) Aplicação de etileno em frutos climatéricos. Estimula a respiração antes da ascensão respiratória, no pré-climatérico. Reação irreversível. Aumenta enzimas ligadas ao amadurecimento. (Kays,1991)")
71
Inibidores Inibidores da ação do etileno
Ligam-se aos receptores nos sítios específicos das células, bloqueando a ação do etileno. Inibidores da biossíntese do etileno Inibem a ação de enzimas (ACC sintase, ACC oxidase), impedindo ou bloqueando a via de síntese e, consequentemente, a produção do etileno. Os inibidores da ação podem proteger os tecidos contra o etileno endógeno e exógeno, causando uma melhor proteção.
, impedindo ou bloqueando a via de síntese e, consequentemente, a produção do etileno. Os inibidores da ação podem proteger os tecidos contra o etileno endógeno e exógeno, causando uma melhor proteção.")
72
(Chitarra & Chitarra, 2005)
")
73
(Chitarra & Chitarra, 2005) 1-MCP = 1-metilciclopropeno
Bloqueador da ação do etileno Retarda o amadurecimento de frutos e senescência de flores cortadas (Chitarra & Chitarra, 2005)
")
74
FATORES DE INFLUÊNCIA NA BIOSSÍNTESE DO ETILENO
Concentração de O2 Concentração de CO2 Variação de temperatura Exposição à luz Condições de estresse ambiental/biológico O dióxido de carbono em elevadas concentrações inibe a ação do etileno sobre os tecidos vegetais e sua formação ocorre a partir da oxidação do Ácido1aminociclopropano carboxílico, pela baixa pressão parcial de oxigênio (VAROQUAUX & WILEY, 1997). Reduzindo o O2 a níveis menores que 8% a produção de etileno é reduzida em frutas e hortaliças. O O2 é necessário para a síntese e ação do etileno, uma vez que sob condições anaeróbicas a conversão do ACC (ácido 1-aminociclopropanocarboxílico) à etileno é inibida, promovendo um acúmulo de ACC no tecido, uma vez que a passagem de metionina a ACC ocorre mesmo na Ausência de O2. Altas concentrações de CO2, por sua vez, pode reduzir, promover ou não ter nenhum efeito na taxa de produção de etileno pelo fruto, dependendo do produto e da concentração a que este é exposto. Em alguns frutos, ocorre um acúmulo de CO2 nos espaços intercelulares e este funciona como um antagonista natural do etileno (Kader, 1992)
. Reduzindo o O2 a níveis menores que 8% a produção de etileno é reduzida em frutas e hortaliças. O O2 é necessário para a síntese e ação do etileno, uma vez que sob condições anaeróbicas a conversão do ACC (ácido 1-aminociclopropanocarboxílico) à etileno é inibida, promovendo um acúmulo de ACC no tecido, uma vez que a passagem de metionina a ACC ocorre mesmo na Ausência de O2. Altas concentrações de CO2, por sua vez, pode reduzir, promover ou não ter nenhum efeito na taxa de produção de etileno pelo fruto, dependendo do produto e da concentração a que este é exposto. Em alguns frutos, ocorre um acúmulo de CO2 nos espaços intercelulares e este funciona como um antagonista natural do etileno (Kader, 1992)")
75
(Chitarra & Chitarra, 2005)
")
77
Efeito do aumento da temperatura na produção de etileno

78
Efeitos desejáveis do etileno
Indução do amadurecimento Uniformização do amadurecimento (ex. banana) Desverdescimento de citrus Estimula a abscisão (facilita a colheita)
 Desverdescimento de citrus. Estimula a abscisão (facilita a colheita)")
79
Efeitos indesejáveis do etileno
Amarelecimento de produtos hortícolas (hortaliças folhosas e flores) Formação de compostos amargos e tóxicos (ex.: isocumarina) Abscisão (folhas e flores) Brotamento (cebola, batata) MÉTODOS PARA EVITAR EFEITOS INDESEJÁVEIS DO ETILENO Não misturar produtos no armazenamento • Evitar fontes liberadoras de etileno • Troca de ar nas câmaras • Absorvedores de etileno (permanganato de K, carvão ativado) • Lâmpada UV ou ozônio (oxidam o etileno) - O2 + UV ---- O3 - O3 + C2H H2O + CO2 + CO + formaldeído
 Formação de compostos amargos e tóxicos (ex.: isocumarina) Abscisão (folhas e flores) Brotamento (cebola, batata) MÉTODOS PARA EVITAR EFEITOS INDESEJÁVEIS DO ETILENO. Não misturar produtos no armazenamento. • Evitar fontes liberadoras de etileno. • Troca de ar nas câmaras. • Absorvedores de etileno (permanganato de. K, carvão ativado) • Lâmpada UV ou ozônio (oxidam o etileno) - O2 + UV ---- O3. - O3 + C2H H2O + CO2 + CO + formaldeído.")
80
HORMÔNIOS DE PLANTAS (FITORMÔNIOS)
Ácido abscíssico (ABA) Estimula a produção de etileno Acelera o processo de senescência
 Estimula a produção de etileno. Acelera o processo de senescência.")
81
HORMÔNIOS DE PLANTAS (FITORMÔNIOS)
Auxinas (ex Ác. 3-indolacético, IAA) Neutralizam os efeitos do etileno e do ácido abscíssico Retardam o processo de senescência
 Neutralizam os efeitos do etileno e do ácido abscíssico. Retardam o processo de senescência.")
82
CITOCININAS (ex. Zeatina) e GIBERELINAS (ex. Giberelina A3)
Retardam o processo de senescência Atuam isoladamente ou em conjunto com as auxinas

83
Outros compostos reguladores
Sinalizadores e/ou precursores de compostos reguladores Aminas bioativas Além dos fitormônios, diferentes grupos de compostos atuam no controle dos processos metabólicos dos vegetais, seja como sinalizadores, seja como elicitores de respostas fisiológicas, ou mesmo como metabólitos intermediários utilizados como precursores para a biossíntese de outros compostos. Do mesmo modo, as estruturas celulares e suas membranas têm o controle dos eventos que conduzem as reações de vida ou morte dos tecidos, evidenciado pelas modificações no código genético, na síntese protéica e na resistência organizacional dessas membranas. As aminas bioativas são bases orgânicas de baixo peso molecular , sintetizadas e degradadas como resultado da atividade metabólica normal de animais, plantas e microrganismos, sendo produzidas pela descarboxilação de aminoácidos, decomposições de fosfolipídeos ou por transaminação de aldeídos, cetonas ou hidrólise de compostos nitrogenados. As frutas e hortaliças apresentam teores variáveis desses compostos em função da espécie, da cultivar, do tipo de tecido, do grau de maturação e/ou de desenvolvimento. (Chitarra & Chitarra, 2005)
")
84
Aula 4 Maturação & Cor; Maturação & Textura; Maturação & Aroma.

85
MATURAÇÃO E COR Após o início do climatérico mudanças graduais de cor ocorrem Fruta Imatura Madura Maçã Verde Amarela/vermelha Banana Amarela Pêra Morango Vermelha Etileno promove degradação da clorofila Evidência outros pigmentos já presentes Estimula a síntese de novo desse pigmentos Carotenóides e antocianinas são os principais A degradação da clorofila durante a maturação de frutas mostra os outros pigmentos existentes bem como acentuam-se suas sínteses

86
Clorofilas As clorofilas são denominadas clorofila a e clorofila b e normalmente se encontram na proporção de 1:3 (clorofila a/ clorofila b) As clorofilas (encontradas nos cloroplastos) têm coloração do azul-esverdeado ao verde-amarelado. São pigmentos receptores da luz (transformam a energia luminosa em química) e desenvolvem a fotossíntese através da fixação de CO2 com liberação de O2 (CHITARRA, 2000). As clorofilas são denominadas clorofila a e clorofila b e se encontram sempre na proporção de 1:3 (clorofila a/ clorofila b) sendo obtidas facilmente por extração com solventes orgânicos, entre os quais o mais indicado é a acetona (BOBBIO & BOBBIO, 2003). Usualmente, com a maturação dos tecidos há degradação da clorofila, tornando visíveis pigmentos pré-existentes e/ou novos pigmentos sintetizados, responsáveis pela coloração característica de cada espécie ou mesmo de cada cultivar (CHITARRA, 2000). Com a evolução da maturação a clorofila é degradada pela ação da enzima clorofilase ou por modificação do pH (devido a liberação de ácidos orgânicos dos vacúolos) e por sistemas oxidativos, com perda do grupo fitol ou de Mg++, formando compostos com coloração variada ou incolores. Na Figura 7a encontra-se a fórmula estrutural da clorofila (CHITARRA, 2000).
 têm coloração do azul-esverdeado ao verde-amarelado. São pigmentos receptores da luz (transformam a energia luminosa em química) e desenvolvem a fotossíntese através da fixação de CO2 com liberação de O2 (CHITARRA, 2000). As clorofilas são denominadas clorofila a e clorofila b e se encontram sempre na proporção de 1:3 (clorofila a/ clorofila b) sendo obtidas facilmente por extração com solventes orgânicos, entre os quais o mais indicado é a acetona (BOBBIO & BOBBIO, 2003). Usualmente, com a maturação dos tecidos há degradação da clorofila, tornando visíveis pigmentos pré-existentes e/ou novos pigmentos sintetizados, responsáveis pela coloração característica de cada espécie ou mesmo de cada cultivar (CHITARRA, 2000). Com a evolução da maturação a clorofila é degradada pela ação da enzima clorofilase ou por modificação do pH (devido a liberação de ácidos orgânicos dos vacúolos) e por sistemas oxidativos, com perda do grupo fitol ou de Mg++, formando compostos com coloração variada ou incolores. Na Figura 7a encontra-se a fórmula estrutural da clorofila (CHITARRA, 2000).")
87
Perda de cor da clorofila
pH (ácidos orgânicos) Substituição do Mg++ por H+ formando feofitina Ação enzimática Clorofilase Peroxidase Lipo-oxigenase Oxidação formando diversos produtos incolores Com a evolução da maturação a clorofila é degradada pela ação da enzima clorofilase ou por modificação do pH (devido a liberação de ácidos orgânicos dos vacúolos) e por sistemas oxidativos, com perda do grupo fitol ou de Mg++, formando compostos com coloração variada ou incolores. Na Figura 7a encontra-se a fórmula estrutural da clorofila (CHITARRA, 2000). A clorofilase é uma esterase que cataliza a clivagem do grupo fitol da molécula clorofila e de seus derivativos livres de Mg++ (feofitinas) formando, respectivamente, clorofilídeos e feofórbidos. A formação de clorofilídeos só ocorre em folhas frescas na fase pós-colheita, quando a enzima é ativada pelo calor. A atividade ótima ocorre em temperaturas superiores a 60°C, sendo inativada a 100°C (CHITARRA, 2000). As clorofilas podem ser alteradas quimicamente por diversos fatores. Por exemplo, o fitol pode ser removido facilmente por álcalis com formação das clorofilidas, compostos de cor verde que possuem praticamente a mesma absorvância das clorofilas, embora sejam bastante mais solúveis em água. No processamento de alimentos a reação mais importante é sem dúvida a facilidade com que o magnésio é substituído por prótons, pela ação de ácidos diluídos, com formação das feofitinas, compostos de cor verde oliva, cor essa que prejudica a aparência de alimentos ricos em clorofilas. In situ, embora os tecidos vegetais sejam ácidos, as clorofilas são bastante estáveis, talvez devido ao fato de se encontrarem associadas a proteínas e lipídios, que teriam ação protetora (BOBBIO & BOBBIO, 2003). As transformações que as clorofilas sofrem estão esquematizadas na Figura 7b.
 Substituição do Mg++ por H+ formando feofitina. Ação enzimática. Clorofilase. Peroxidase. Lipo-oxigenase. Oxidação formando diversos. produtos incolores. Com a evolução da maturação a clorofila é degradada pela ação da enzima clorofilase ou por modificação do pH (devido a liberação de ácidos orgânicos dos vacúolos) e por sistemas oxidativos, com perda do grupo fitol ou de Mg++, formando compostos com coloração variada ou incolores. Na Figura 7a encontra-se a fórmula estrutural da clorofila (CHITARRA, 2000). A clorofilase é uma esterase que cataliza a clivagem do grupo fitol da molécula clorofila e de seus derivativos livres de Mg++ (feofitinas) formando, respectivamente, clorofilídeos e feofórbidos. A formação de clorofilídeos só ocorre em folhas frescas na fase pós-colheita, quando a enzima é ativada pelo calor. A atividade ótima ocorre em temperaturas superiores a 60°C, sendo inativada a 100°C (CHITARRA, 2000). As clorofilas podem ser alteradas quimicamente por diversos fatores. Por exemplo, o fitol pode ser removido facilmente por álcalis com formação das clorofilidas, compostos de cor verde que possuem praticamente a mesma absorvância das clorofilas, embora sejam bastante mais solúveis em água. No processamento de alimentos a reação mais importante é sem dúvida a facilidade com que o magnésio é substituído por prótons, pela ação de ácidos diluídos, com formação das feofitinas, compostos de cor verde oliva, cor essa que prejudica a aparência de alimentos ricos em clorofilas. In situ, embora os tecidos vegetais sejam ácidos, as clorofilas são bastante estáveis, talvez devido ao fato de se encontrarem associadas a proteínas e lipídios, que teriam ação protetora (BOBBIO & BOBBIO, 2003). As transformações que as clorofilas sofrem estão esquematizadas na Figura 7b.")
88
Esquema da degradação da clorofila por diferentes agentes (CHITARRA, 2000)
")
89
Carotenóides Os carotenóides são corantes naturais responsáveis pelo espectro de cores que varia do amarelo ao vermelho; Químicamente são substâncias tetraterpênicas (C40) formadas por 8 unidades de isopreno (C5). Cerca de 600 estruturas de carotenóides foram identificadas; São divididos em carotenos, compostos constituídos apenas por carbono e hidrogênio e seus derivados oxigenados, as xantofilas; Localizam-se nos cromoplastos e nos cloroplastos; Fotossíntese: absorção de luz e fotoproteção. Os carotenoides de animais provalvelmente são produtos resultantes de mudanças metabolicas, geralmente oxidativas, da ingestão de carotenoides existentes em vegetais Quimicamente os carotenóides são substâncias tetraterpênicas, formadas por oito unidades de isopropeno, ligadas covalentemente cabeça-cauda e cauda-cauda, para criar uma molécula simétrica. A ligação isoprênica sofre reversão na parte central da molécula, e dessa maneira os dois grupos metílicos centrais ficam separados por quatro carbonos, resultando numa molécula simétrica. Algumas estruturas contêm tb grupamentos cíclicos nas extremidades como o beta-caroteno Os carotenóides são divididos em carotenos, compostos constituídos apenas por carbono e hidrogênio, e seus derivados oxigenados, as xantofilas. Possuem cor intensa, que varia do amarelo ao vermelho, mudando para azul por reação com ácido sulfúrico ou tricloreto de antimônio (BOBBIO & BOBBIO, 2003). Os pigmentos carotenóides localizam-se nos cromoplastos e também nos cloroplastos, associados com a clorofila. Têm como funções a proteção da clorofila e do aparelho fotossintético contra a fotodegradação (protegendo a molécula da clorofila), bem como a absorção de luz em comprimento de onda diferente do da clorofila, aumentando o potencial energético do sistema (CHITARRA, 2000).
 formadas por 8 unidades de isopreno (C5). Cerca de 600 estruturas de carotenóides foram identificadas; São divididos em carotenos, compostos constituídos apenas por carbono e hidrogênio e seus derivados oxigenados, as xantofilas; Localizam-se nos cromoplastos e nos cloroplastos; Fotossíntese: absorção de luz e fotoproteção. Os carotenoides de animais provalvelmente são produtos resultantes de mudanças metabolicas, geralmente oxidativas, da ingestão de carotenoides existentes em vegetais. Quimicamente os carotenóides são substâncias tetraterpênicas, formadas por oito unidades de isopropeno, ligadas covalentemente cabeça-cauda e cauda-cauda, para criar uma molécula simétrica. A ligação isoprênica sofre reversão na parte central da molécula, e dessa maneira os dois grupos metílicos centrais ficam separados por quatro carbonos, resultando numa molécula simétrica. Algumas estruturas contêm tb grupamentos cíclicos nas extremidades como o beta-caroteno. Os carotenóides são divididos em carotenos, compostos constituídos apenas por carbono e hidrogênio, e seus derivados oxigenados, as xantofilas. Possuem cor intensa, que varia do amarelo ao vermelho, mudando para azul por reação com ácido sulfúrico ou tricloreto de antimônio (BOBBIO & BOBBIO, 2003). Os pigmentos carotenóides localizam-se nos cromoplastos e também nos cloroplastos, associados com a clorofila. Têm como funções a proteção da clorofila e do aparelho fotossintético contra a fotodegradação (protegendo a molécula da clorofila), bem como a absorção de luz em comprimento de onda diferente do da clorofila, aumentando o potencial energético do sistema (CHITARRA, 2000).")
90
Biossíntese dos carotenóides
São formados a partir do Isopentenil Pirofosfato (IPP) e Dimetilalil Difosfato (DMAPP), no cloroplasto (e cromoplasto). O IPP é sintetizado na rota do ácido mevalônico (MVA) O DMAPP é sintetizado na rota do metil eritritol fosfato Fitoeno é a estrutura básica para formação de carotenos A coloração vai sendo adquirida a partir de desaturações produzindo um sistema de duplas ligações conjugadas Falar do sistema de duplas conjugadas pelo meons 7 duplas conjugadas Os terpenos são biossintetizados a partir de metabólitos primários por duas rotas diferentes: a rota do ácido mevalônico, onde três moléculas de acetil CoA são ligadas, a partir de urna série etapas da rota, para formar o ácido mevalônico. Esse importante intermediário de seis carbonos e então pirofosforilado, descarboxilado e desidratado para produzir isopentenil difosfato (IPP) e a rota do metileritritol fosfato, onde o gliceraldeído-3-fosfato e dois átomos de carbono derivados do piruvato se combinam para formar o dimetilalil difosfato (DMAPP) que, eventualmente, e convertido em IPP (Taiz & Zeiger, 2004) O isopentenil difosfato e seu isômero, dimetilalil difosfato são unidades pentacarbonadas ativas na biossíntese dos terpenos que se unem para formar moléculas maiores, entre elas o geranilgeranil difosfato (GGPP), um composto de 20 carbonos (Figura 2) (Taiz & Zeiger, 2004).
 e Dimetilalil Difosfato (DMAPP), no cloroplasto (e cromoplasto). O IPP é sintetizado na rota do ácido mevalônico (MVA) O DMAPP é sintetizado na rota do metil eritritol fosfato. Fitoeno é a estrutura básica para formação de carotenos. A coloração vai sendo adquirida a partir de desaturações produzindo um sistema de duplas ligações conjugadas. Falar do sistema de duplas conjugadas pelo meons 7 duplas conjugadas. Os terpenos são biossintetizados a partir de metabólitos primários por duas rotas diferentes: a rota do ácido mevalônico, onde três moléculas de acetil CoA são ligadas, a partir de urna série etapas da rota, para formar o ácido mevalônico. Esse importante intermediário de seis carbonos e então pirofosforilado, descarboxilado e desidratado para produzir isopentenil difosfato (IPP) e a rota do metileritritol fosfato, onde o gliceraldeído-3-fosfato e dois átomos de carbono derivados do piruvato se combinam para formar o dimetilalil difosfato (DMAPP) que, eventualmente, e convertido em IPP (Taiz & Zeiger, 2004) O isopentenil difosfato e seu isômero, dimetilalil difosfato são unidades pentacarbonadas ativas na biossíntese dos terpenos que se unem para formar moléculas maiores, entre elas o geranilgeranil difosfato (GGPP), um composto de 20 carbonos (Figura 2) (Taiz & Zeiger, 2004).")
91
Biossíntese dos Terpenos
Os carotenóides são formados em microorganismos e plantas, mas não nos mamíferos O licopeno pode sofrer ciclização dando origem ao beta e alfa caroteno, seguida de oxidação gerando as xantofilas Terpenos Taiz & Zeiger, 2002

92
Biossíntese dos demais Carotenóides
O licopeno pode sofrer ciclização dando origem ao beta e alfa caroteno, seguida de oxidação gerando as xantofilas Kopsell & Kopssel, 2002

93
MUDANÇAS NA COMPOSIÇÃO EM CAROTENÓIDES DA CASA DE CAQUI NA PÓS-COLHEITA
Ripening stage Harvet-ripe Intermediate Fully ripe Total carotenoids (g/g fr. Wt) 128.0 366.0 491.0 Carotenoid pattern (% of total carotenoids) Phytofluene - 0.4 -Carotene 1.6 1.2 1.0 - Carotene 9.4 7.6 6.7 Mutatochrome 0.7 - Carotene Lycopene 1.1 0.5 8.2 -Cryptoxanthin 29.2 50.0 48.2 Cryptoxanthin 5,6-epoxide 0.9 1.9 Cryptoflavin 2.1 2.9 Lutein 12.4 5.5 4.1 Zeaxanthin 9.3 9.7 5.9 Mutatoxanthin 0.8 4.7 1.8 Isolutein 0.3 trans-Antheraxanthin 5.4 2.0 4.8 cis-Antheraxanthin 6.2 2.2 2.3 Luteoxanthin 1.7 trans-violaxanthin 6.9 3.7 3.8 cis-violaxanthin 1.5 Neoxanthin 7.2 5.2 From Ebert and Gross (1985).
 Carotenoid pattern (% of total carotenoids) Phytofluene -Carotene - Carotene Mutatochrome - Carotene. Lycopene -Cryptoxanthin Cryptoxanthin 5,6-epoxide Cryptoflavin Lutein Zeaxanthin Mutatoxanthin Isolutein trans-Antheraxanthin cis-Antheraxanthin Luteoxanthin trans-violaxanthin cis-violaxanthin Neoxanthin From Ebert and Gross (1985).")
94
Carotenóides Com a degradação da clorofila, os carotenóides previamente presentes nos tecidos tornam-se visíveis ou podem também ser sintetizados com o avanço da maturação dos frutos. Sistema de classificação de bananas de acordo com seu grau de maturação (VILAS BOAS et al., 2001). Os pigmentos carotenóides podem já estar presentes, tomando-se visíveis com a degradação da clorofila; ou podem ser sintetizados, simultaneamente, com a degradação substancia. Em tomates, ha uma intensa degradacão de clorofila durante o amadurecimento, com síntese gradual do licopeno Em bananas, a degradacão da clorofila é o principal evento, ao passo que a síntese de outros pigmentos e realizada em níveis relativamente baixos. De modo oposto, em algumas cultivares de maça, as modificacões da cor da casca são devido, preferencialmente, a síntese de carotenos e de antocianinas do que uma grande degradacão da clorofila. Modificação nos pigmentos do tomate durante a maturação A) Verde-maturo; B) Verde- amarelo; C) Amarelo-laranja com alguns traços verdes; D) Laranja-amarelo,sem traços verdes; E) Laranja-vermelho; F) Vermelho (Chitarra & Chitarra, 2005)
. Os pigmentos carotenóides podem já estar presentes, tomando-se visíveis com a degradação da clorofila; ou podem ser sintetizados, simultaneamente, com a degradação substancia. Em tomates, ha uma intensa degradacão de clorofila durante o amadurecimento, com síntese gradual do licopeno Em bananas, a degradacão da clorofila é o principal evento, ao passo que a síntese de outros pigmentos e realizada em níveis relativamente baixos. De modo oposto, em algumas cultivares de maça, as modificacões da cor da casca são devido, preferencialmente, a síntese de carotenos e de antocianinas do que uma grande degradacão da clorofila. Modificação nos pigmentos do tomate durante a maturação A) Verde-maturo; B) Verde- amarelo; C) Amarelo-laranja com alguns traços verdes; D) Laranja-amarelo,sem traços verdes; E) Laranja-vermelho; F) Vermelho (Chitarra & Chitarra, 2005)")
95
ANTOCIANINAS NA MATURAÇÃO
Responsáveis pelas cores vermelha, violeta e azul Presente nas folhas, frutos, folhas e raízes São um subgrupo de flavonóides cuja estrutura é baseada no cátion flavílio, geralmente conjugadas com açúcares simples no C 3 (e C 5) do anel C.
 do anel C.")
96
As antocianidinas são as agliconas. Exemplos:
Pelargonidina, cianidina, delfinidina e malvidina

97
Algumas Fontes de Antocianidinas
Fruta Cianidina Cereja preta, ruibarbo Cianidina, delfinidina Amora preta Cianidina, peonidina Cereja Cianidina, pelargonidina Morango A fenilalanina amônia-liase destaca-se como uma enzima chave e regulatória da rota de biossíntese dos fenilpropanóides e seus derivados, catalisando a transformação, por desaminação, do aminoácido Lfenilalanina, em ácido trans-cinâmico, sendo este o primeiro passo para a biossíntese dos fenólicos vegetais (Cheng et al., 2001). Do ponto de vista fisiológico, os taninos são considerados metabólitos secundários, originados diretamente do metabolismo do carbono (Koricheva et al., 1998; Heyworth et al., 1998). Nas plantas, a alocação de carbono para a síntese de metabólitos secundários é determinada tanto pela disponibilidade de recursos edafoclimáticos (água, nutrientes, luz, concentração de CO2), como por estresses específicos (seca, poluentes atmosféricos, radiação ultra-violeta, temperatura) (Koricheva et al., 1998; Lavola, 1998; Kouki & Manetas, 2002). Assim, vários fatores que afetam o crescimento, fotossíntese e outros aspectos do metabolismo primário, também causam efeitos no metabolismo secundário (Lavola et al., 2000). Entretanto, segundo Lavola (1998), deve-se considerar que quando associados, estes fatores podem afetar diferentemente os padrões morfológicos e bioquímicos das plantas.
. Do ponto de vista fisiológico, os taninos são considerados metabólitos secundários, originados diretamente do metabolismo do carbono (Koricheva et al., 1998; Heyworth et al., 1998). Nas plantas, a alocação de carbono para a síntese de metabólitos secundários é determinada tanto pela disponibilidade de recursos edafoclimáticos (água, nutrientes, luz, concentração de CO2), como por estresses específicos (seca, poluentes atmosféricos, radiação ultra-violeta, temperatura) (Koricheva et al., 1998; Lavola, 1998; Kouki & Manetas, 2002). Assim, vários fatores que afetam o crescimento, fotossíntese e outros aspectos do metabolismo primário, também causam efeitos no metabolismo secundário (Lavola et al., 2000). Entretanto, segundo Lavola (1998), deve-se considerar que quando associados, estes fatores podem afetar diferentemente os padrões morfológicos e bioquímicos das plantas.")
98
Biosíntese de Flavonoides em Plantas
Fenil Alanina Amonia Liase Ac. p-cumárico De acordo com Kouki & Manetas (2002), a disponibilidade de recursos também promove efeitos regulatórios na atividade de enzimas chaves da rota biossintética destes metabólitos. Janas et al. (2000) ressaltam que a atividade da fenilalanina amônia-liase e o acúmulo de fenólicos ocorrem como conseqüência de estresses bióticos, como infecção por microorganismos, ataque de insetos e herbívoros (Rivero et al., 2001) e estresses abióticos como temperatura (Solecka et al., 1999; Janas et al., 2000), luz (Lavola et al., 2000), disponibilidade de água (Chakraborty et al., 2001), ferimentos (Saltveit, 2000) e níveis de CO2 atmosférico (Hartley et al., 2000; Assis et al., 2001). Solecka & Kacperska (1995) relatam que estes estresses podem modificar tanto a síntese da enzima, como a sua atividade, em uma ampla variedade de plantas. Eskin, 1990
, a disponibilidade de recursos também promove efeitos regulatórios na atividade de enzimas chaves da rota biossintética destes metabólitos. Janas et al. (2000) ressaltam que a atividade da fenilalanina amônia-liase e o acúmulo de fenólicos ocorrem como conseqüência de estresses bióticos, como infecção por microorganismos, ataque de insetos e herbívoros (Rivero et al., 2001) e estresses abióticos como temperatura (Solecka et al., 1999; Janas et al., 2000), luz (Lavola et al., 2000), disponibilidade de água (Chakraborty et al., 2001), ferimentos (Saltveit, 2000) e níveis de CO2 atmosférico (Hartley et al., 2000; Assis et al., 2001). Solecka & Kacperska (1995) relatam que estes estresses podem modificar tanto a síntese da enzima, como a sua atividade, em uma ampla variedade de plantas. Eskin,")
99
As antocianinas são anfóteras, em diferentes pH esses pigmentos se encontram em diferentes formas e apresentam cores diversas. Estrutura química da antocianina em diferentes pH (BOBBIO & BOBBIO, 2003)
")
100
MODIFICAÇÃO DA TEXTURA DURANTE A MATURAÇÃO
Degradação de componentes estruturais da parede celular Amaciamento dos tecidos Celulose Pectina: galacturonanas, com diferentes graus de residuos metilados Hemiceluloses Proteínas estruturais Lignina: polímeros complexos de derivados do fenil propano Entretanto, as lesões provocadas durante o processamento promovem a descompartimentalização celular e possibilitam o contato de enzimas e substratos, que originam modificações bioquímicas, como escurecimento, formação de odores desagradáveis e perda da textura original. Além disso, o descascamento e o corte de frutas e vegetais favorecem a colonização dos tecidos vegetais por microrganismos deterioradores e patogênicos. A degradação da textura, deve-se a ação de proteases e enzimas pectinolíticas dos compostos da parede celular para ao interior do produto (VAROQUAUX & WILEY, 1997). Altas concentrações de CO2 inibem a degradação de textura em morangos, enquanto que não altera o amolecimento de rodelas de kiwi, sugerindo que a concentração gasosa não exerce efeito sobre as enzimas proteolíticas e pectinolíticas (VAROQUAUX & WILEY, 1997).
. Altas concentrações de CO2 inibem a degradação de textura em morangos, enquanto que não altera o amolecimento de rodelas de kiwi, sugerindo que a concentração gasosa não exerce efeito sobre as enzimas proteolíticas e pectinolíticas (VAROQUAUX & WILEY, 1997).")
101
Parede celular Componentes: Celulose, hemicelulose, pectinas, lignina e proteínas estruturais Parede celular primária: As microfibrilas de celulose são revestidas com hemicelulose (como o xiloglucano), que podem também ligar as microfibrilas entre si. As pectinas formam um gel de matriz de ligação, talvez interagindo com proteínas estruturais Hemiceluloses e celulose representam mais de 50% do peso seco de resíduos agrícolas (7), eles podem ser convertidos em açúcares solúveis, quer por hidrólise ácida ou enzimática, de modo a que possam ser utilizados como uma abundante e barata fonte de energia renovável no mundo. Dentre elas salientam-se a fenilalanina amônialiase (FAL), enzima chave na síntese de lignina (Hahlbrock e Scheel, 1989); a peroxidase aniônica, localizada na parede celular, responsável pela biossíntese de lignina, baseado na sua afinidade com os álcoois cinamil, (Lagrimini, 1987) e a polifenoloxidase, responsável por muitas reações de escurecimento durante o manuseio, armazenamento e processamento de frutos (Mayer e Harel, 1979; Bower e Cutting, 1988), assim como por reações oxidativas associadas com o escurecimento e descoloração indesejáveis em muitos tecidos de frutos e hortaliças frescos injuriados (Luh e Phithakpol, 1972; Siddiq, Sinha e Cash, 1992). (artigo pêssego Chitarra) Taiz & Zeiger, 2002
, que podem também ligar as microfibrilas entre si. As pectinas formam um gel de matriz de ligação, talvez interagindo com proteínas estruturais. Hemiceluloses e celulose representam mais de 50% do peso seco de resíduos agrícolas (7), eles podem ser convertidos em açúcares solúveis, quer por hidrólise ácida ou enzimática, de modo a que possam ser utilizados como uma abundante e barata fonte de energia renovável no mundo. Dentre elas salientam-se a fenilalanina amônialiase (FAL), enzima chave na síntese de lignina (Hahlbrock e Scheel, 1989); a peroxidase aniônica, localizada na parede celular, responsável pela biossíntese de lignina, baseado na sua afinidade com os álcoois cinamil, (Lagrimini, 1987) e a polifenoloxidase, responsável por muitas reações de escurecimento durante o manuseio, armazenamento e processamento de frutos (Mayer e Harel, 1979; Bower e Cutting, 1988), assim como por reações oxidativas associadas com o escurecimento e descoloração indesejáveis em muitos tecidos de frutos e hortaliças frescos injuriados (Luh e Phithakpol, 1972; Siddiq, Sinha e Cash, 1992). (artigo pêssego Chitarra) Taiz & Zeiger,")
102
Celulose (mecanismo de degradação na maturação não é muito claro)
Celulose insolúvel C1-celulase Derivados solúveis Cx-celulase Celobiose Celobiase (-1,4-glicosidase) Glicose
 Glicose.")
103
-Galactosidase (> atividade na maturação)
-(1→4)-galactana galactose Degradação de Pectinas atividade Poligalacturonases (PG) atividade Pectina metil esterases (PME) Pectinas Insolúveis Pectinas solúveis
-galactana galactose. Degradação de Pectinas. atividade Poligalacturonases (PG) atividade Pectina metil esterases (PME) Pectinas Insolúveis. Pectinas solúveis.")
104
PME = pectinametilesterase PG = poligalacturonase (CHITARRA, 2000).
Durante o crescimento da fruta, a protopectina mantém uma forte coesão das células e, por conseqüência, a fruta apresenta-se extremamente firme antes da sua maturação. No transcorrer da maturação, após a fruta atingir o tamanho máximo, o cálcio é desprendido da protopectina pela ação da enzima protopectinase, iniciando a solubilização da pectina. Outras duas enzimas chave atuam na modificação da textura: a poligalacturonase (PG) e a pectinametilesterase (PME). A PG atua na despolimerização ou encurtamento da cadeia do ácido poligalacturônico, enquanto que a PME promove a desesterificação ou remoção dos grupos metílicos ou acetil da cadeia. Com a atividade destas enzimas, a pectina passa da forma insolúvel para a forma solúvel, observando-se um marcante amaciamento da polpa das frutas (KLUGE, 2002). As pectinas encontram-se nos frutos em diferentes formas, caracterizadas por diferentes solubilidades dependendo do estádio evolutivo do desenvolvimento. A protopectina é uma forma insolúvel em água, esta por hidrólise parcial produz ácidos pectínicos (esterificados com grupos metílicos) ou ácidos pécticos (sem esterificação) também chamados de pectinas solúveis (Figura 11). Estrutura química e mecanismo de solubilização das pectinas pela ação das pectinases PME = pectinametilesterase PG = poligalacturonase (CHITARRA, 2000).
 e a pectinametilesterase (PME). A PG atua na despolimerização ou encurtamento da cadeia do ácido poligalacturônico, enquanto que a PME promove a desesterificação ou remoção dos grupos metílicos ou acetil da cadeia. Com a atividade destas enzimas, a pectina passa da forma insolúvel para a forma solúvel, observando-se um marcante amaciamento da polpa das frutas (KLUGE, 2002). As pectinas encontram-se nos frutos em diferentes formas, caracterizadas por diferentes solubilidades dependendo do estádio evolutivo do desenvolvimento. A protopectina é uma forma insolúvel em água, esta por hidrólise parcial produz ácidos pectínicos (esterificados com grupos metílicos) ou ácidos pécticos (sem esterificação) também chamados de pectinas solúveis (Figura 11). Estrutura química e mecanismo de solubilização das pectinas pela ação das pectinases. PME = pectinametilesterase. PG = poligalacturonase (CHITARRA, 2000).")
105
MODIFICAÇÕES DA PAREDE CELULAR DE GOIABA DURANTE A MATURAÇÃO

106
Melo & Vilas Boas (2007)
")
107
MATURAÇÃO E AROMA No amadurecimento de frutas a biodegradação resulta na formação de alguns compostos aromáticos; Esses compostos vêm de várias rotas diferentes, como: metabolismo de ácidos graxos; metabolismo de aminoácidos; metabolismo de fenólicos; Metabolismo de terpenóides, Os compostos aromáticos vêm de várias rotas diferentes, como do metabolismo de ácidos graxos, aminoácidos, fenólicos e terpenóides, os quais são substratos para determinadas enzimas, formando produtos que serão novamente substrato para outras enzimas em uma rota diferente. O metabolismo de lipídios é, provavelmente, o caminho para a formação de aldeídos, álcoois, ésteres e ácidos, os quais são grupos importantes nas características aromáticas de diversos frutos. A ação da enzima lipoxigenase é o primeiro passo na oxidação dos ácidos linolênico (18:2) e linoléico (18:3), os quais são clivados pela enzima hidroperóxido liase (HPL) para formar aldeídos, ésteres e ácidos (Knee, 2002).
 e linoléico (18:3), os quais são clivados pela enzima hidroperóxido liase (HPL) para formar aldeídos, ésteres e ácidos (Knee, 2002).")
108
MATURAÇÃO E AROMA (Chitarra & Chitarra, 2005)
")
109
Formação de compostos voláteis em banana
Wyllie & Fellman, 2000

110
MATURAÇÃO E AROMA O metabolismo de lipídios é, provavelmente, o caminho para a formação de aldeídos, álcoois, ésteres e ácidos, os quais são grupos importantes nas características aromáticas de diversos frutos. A ação da enzima lipoxigenase é o primeiro passo na oxidação dos ácidos linolênico (18:2) e linoléico (18:3), que leva a formação de aldeídos, ésteres e ácidos. Os compostos aromáticos vêm de várias rotas diferentes, como do metabolismo de ácidos graxos, aminoácidos, fenólicos e terpenóides, os quais são substratos para determinadas enzimas, formando produtos que serão novamente substrato para outras enzimas em uma rota diferente. O metabolismo de lipídios é, provavelmente, o caminho para a formação de aldeídos, álcoois, ésteres e ácidos, os quais são grupos importantes nas características aromáticas de diversos frutos. A ação da enzima lipoxigenase é o primeiro passo na oxidação dos ácidos linolênico (18:2) e linoléico (18:3), os quais são clivados pela enzima hidroperóxido liase (HPL) para formar aldeídos, ésteres e ácidos (Knee, 2002).
 e linoléico (18:3), que leva a formação de aldeídos, ésteres e ácidos. Os compostos aromáticos vêm de várias rotas diferentes, como do metabolismo de ácidos graxos, aminoácidos, fenólicos e terpenóides, os quais são substratos para determinadas enzimas, formando produtos que serão novamente substrato para outras enzimas em uma rota diferente. O metabolismo de lipídios é, provavelmente, o caminho para a formação de aldeídos, álcoois, ésteres e ácidos, os quais são grupos importantes nas características aromáticas de diversos frutos. A ação da enzima lipoxigenase é o primeiro passo na. oxidação dos ácidos linolênico (18:2) e linoléico (18:3), os quais são clivados pela enzima hidroperóxido liase (HPL) para formar aldeídos, ésteres e ácidos (Knee, 2002).")
111
MODELO DE FORMAÇÃO DE COMPONENTES DE AROMA EM FRUTOS DURANTE O AMADURECIMENTO A PARTIR DE AGPI
Eskin, 1990

112
Compostos como β-ionona, α-ionona, diidroactinidiolida, damascenol e β-ciclocitral são alguns dos voláteis derivados de carotenóides. β-caroteno β-ionona os compostos voláteis podem contribuir para o aroma e sabor típico de cada espécie e/ou cultivar. Compostos de aroma derivados de carotenóides têm atraído a atenção de químicos e aromistas há algum tempo, o que resultou no isolamento e elucidação da estrutura química de muitos dos compostos de aroma derivados dos carotenóides obtidos por extração de plantas. Compostos como β-ionona, α-ionona, diidroactinidiolida, damascenol e β-ciclocitral são alguns dos voláteis derivados de carotenóides que, nas plantas, têm a função de antifungicida, afastar herbívoros e até mesmo atrair agentes polinizadores Compostos de aromas advindos de carotenóides podem ser produzidos via enzimática ou não enzimática. A clivagem não enzimática inclui fotoxigenação, (auto) oxidação e degradação térmica. A biodegradação é catalisada por sistemas de dioxigenases. Em geral, três passos são necessários para se gerar um composto de aroma a partir de um carotenóide: clivagem inicial por dioxigenase; subseqüente transformação enzimática dos produtos iniciais de clivagem gerando intermediários polares (precursores de aromas) e conversão catalisada por ácidos de precursores não voláteis na forma ativa dos compostos de aroma. Um exemplo desta reação é a formação de β-damascenona a partir de neoxantina (Figura 1). A primeira clivagem oxidativa do precursor produz uma cetona (denominada “grasshopper ketone”) que é reduzida enzimaticamente e convertida ao produto de interesse. Por outro lado, o β-caroteno é convertido a β-ionona por meio de apenas uma clivagem (Figura 2). Formação de β-ionona a partir de β-caroteno (Uenojo et al. 2007) Esquema geral para formação de compostos de aroma por meio da clivagem de carotenóides e exemplo de formação de β-damascenona a partir de neoxantina (Uenojo et al. 2007)
 oxidação e degradação térmica. A biodegradação é catalisada por sistemas de dioxigenases. Em geral, três passos são necessários para se gerar um composto de aroma a partir de um carotenóide: clivagem inicial por dioxigenase; subseqüente transformação enzimática dos produtos iniciais de clivagem gerando intermediários polares (precursores de aromas) e conversão catalisada por ácidos de precursores não voláteis na forma ativa dos compostos de aroma. Um exemplo desta reação é a formação de β-damascenona a partir de neoxantina (Figura 1). A primeira clivagem oxidativa do precursor produz uma cetona (denominada grasshopper ketone ) que é reduzida enzimaticamente e convertida ao produto de interesse. Por outro lado, o β-caroteno é convertido a β-ionona por meio de apenas uma clivagem (Figura 2). Formação de β-ionona a partir de β-caroteno (Uenojo et al. 2007) Esquema geral para formação de compostos de aroma por meio da clivagem de carotenóides e exemplo de formação de β-damascenona a partir de neoxantina (Uenojo et al. 2007)")
113
BIOSÍNTESE DO ÁCIDO MEVALÔNICO
MVA Eskin, 1990

114
BIOSÍNTESE DO ISOPENTENIL PIROFOSFATO IPP
Eskin, 1990

115
Mecanismo de biossíntese do fitoeno
Eskin, 1990

116
Biossíntese do β-caroteno e do α –caroteno pela ação da licopeno ciclase

Apresentações semelhantes
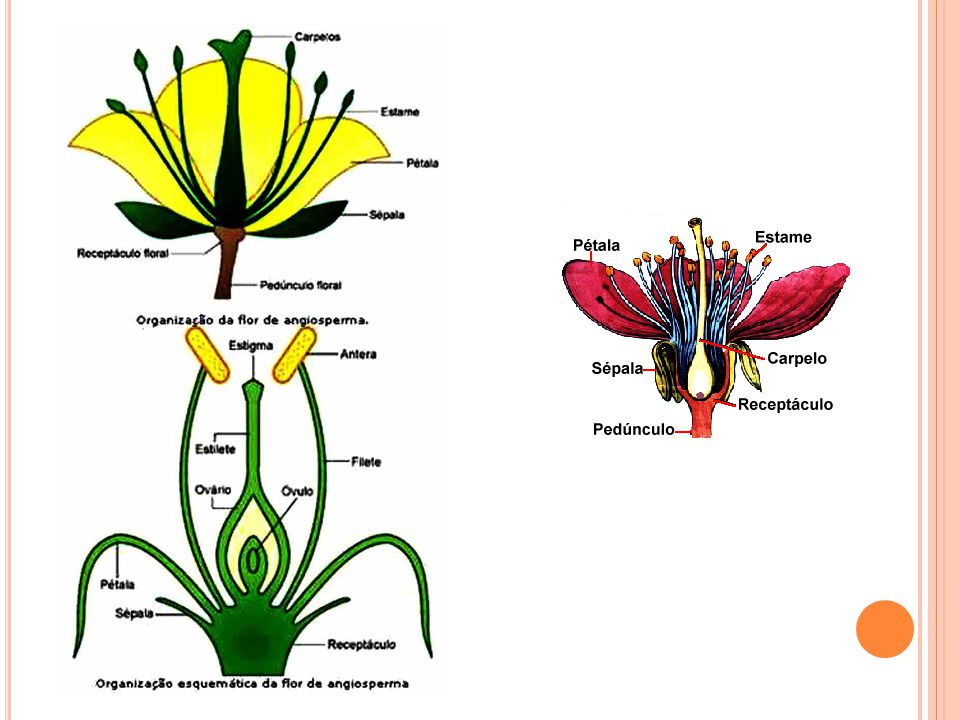

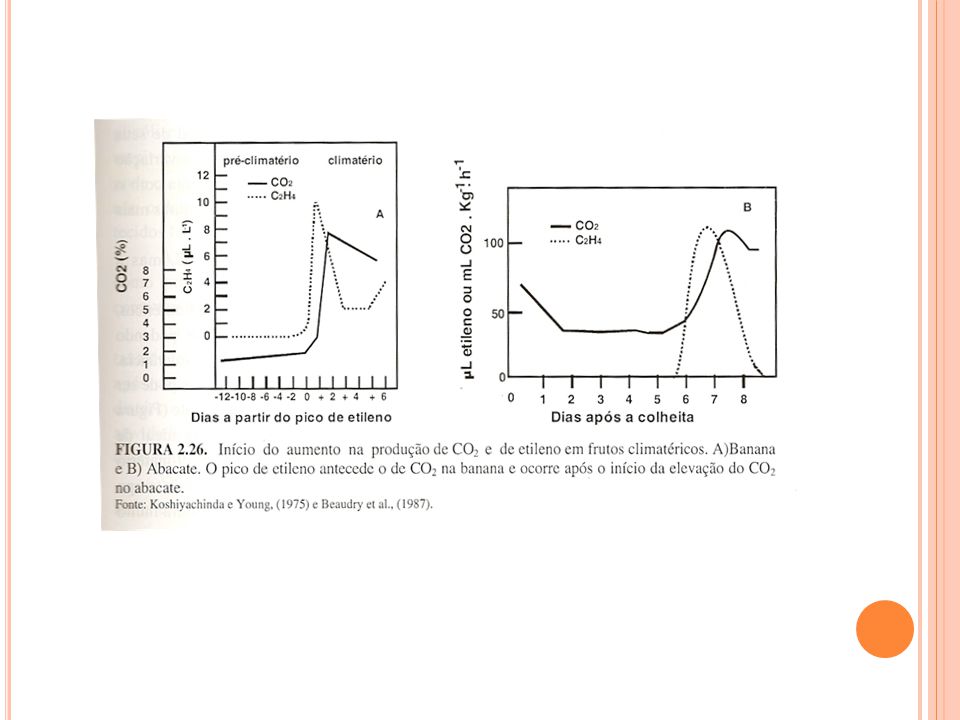

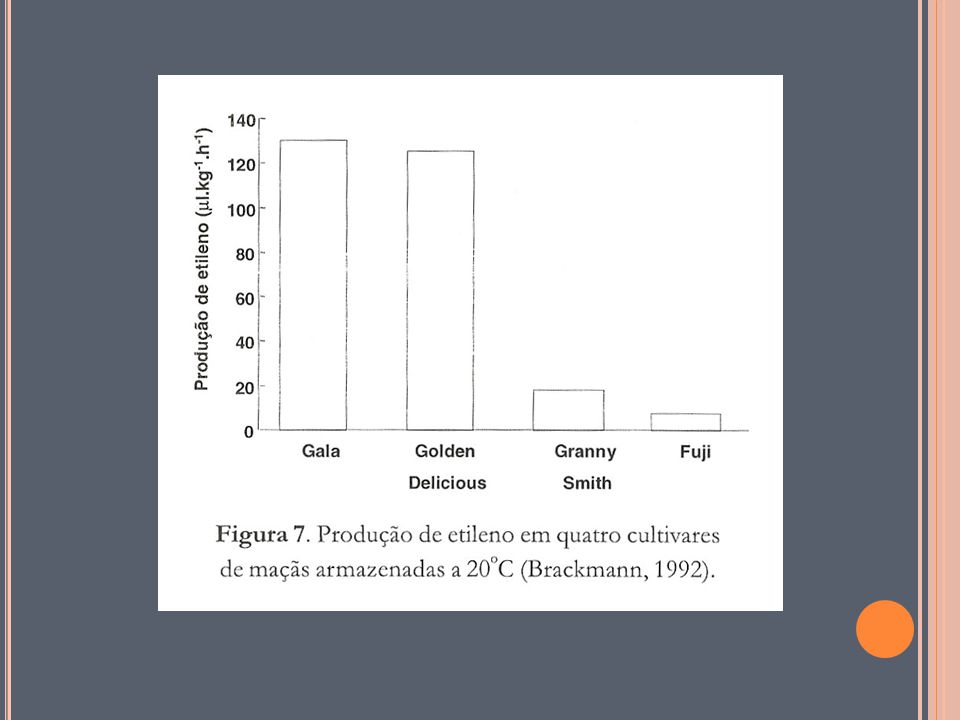

















 NUCLEOTÍDEO = adenosina monofosfato (AMP)Adenosina difosfato (ADP) Adenosina trifosfato (ATP) Adenina Fosfato.>")

