Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Ciclo do Ácido Cítrico Thais Larissa Araújo O. Silva.

2
O manômetro de Otto Warburg - 1920
Era usado para medir a produção de CO2. Permitia determinar a estequiometria das reações. Coloca o substrato Papel de filtro com KOH Tecido Meio de cultura Fluido com cor KOH + CO KHCO3 CO2 KHCO3 Pressão no frasco Leitura do líquido Glicose + 6O CO H2O

3
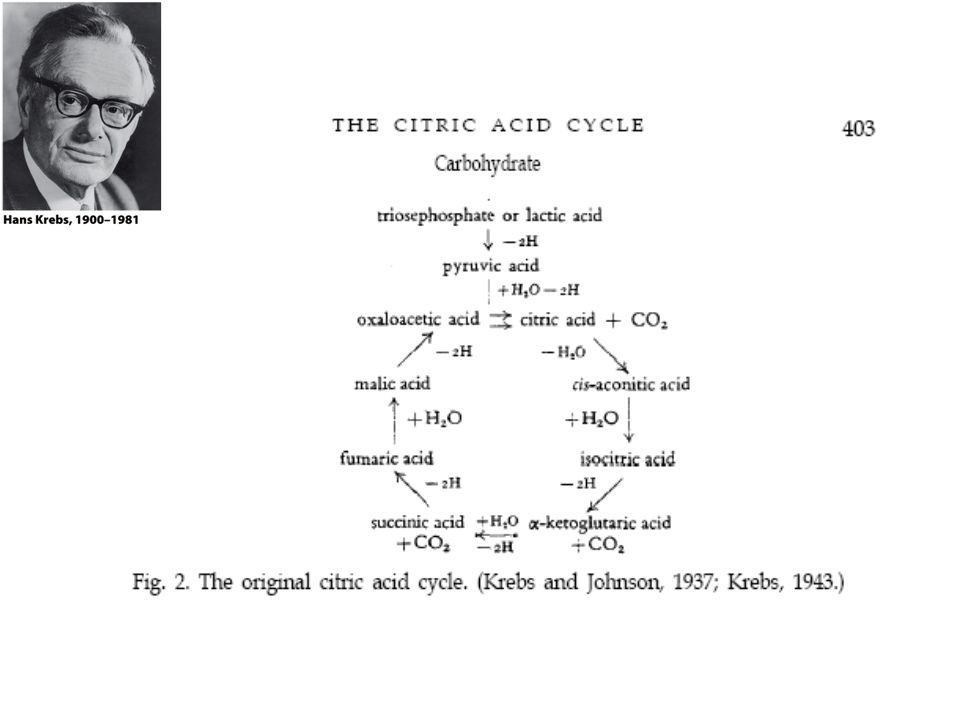
Os experimentos de Hans Krebs
Observando os dados disponíveis na época Krebs destaca os trabalhos de Thumberg entre 1906 e 1920 usando tecidos musculares. Ele testou a oxidação de cerca de 60 substâncias orgânicas e descobriu que a forma ionizada de vários ácidos como o lactato (1 carboxila), succinato, fumarato, malato, citrato (2 carboxilas), eram rapidamente oxidadas. Krebs então testa outros ácidos dicarboxílicos. Em 1935 descobre que um deles, o α-cetoglutarato, com 5 carbonos, assim como nos experimentos de Szent-Györgyi, aceleravam a produção de CO2 e não eram consumidos na reação. Em 1937 Krebs testa ácidos tricarboxílicos como citrato, isocitrato e aconitato, agora com 6 carbonos, e observa que a produção de CO2 também era estimulada e esses intermediários não eram consumidos.
, succinato, fumarato, malato, citrato (2 carboxilas), eram rapidamente oxidadas. Krebs então testa outros ácidos dicarboxílicos. Em 1935 descobre que um deles, o α-cetoglutarato, com 5 carbonos, assim como nos experimentos de Szent-Györgyi, aceleravam a produção de CO2 e não eram consumidos na reação. Em 1937 Krebs testa ácidos tricarboxílicos como citrato, isocitrato e aconitato, agora com 6 carbonos, e observa que a produção de CO2 também era estimulada e esses intermediários não eram consumidos.")
4
Krebs observou nos trabalhos de Shiffield em 1937 que a formação de citrato (C6) ocorria rapidamente após a adição de oxaloacetato (C4) em diversos tecidos. Concluiu então que a formação desse composto de 6 carbonos poderia se originar da ligação de um produto de 4 carbonos (oxaloacetato) mais dois carbonos vindos provavelmente da degradação da glicose. Juntando as seguintes informações: 1- ácidos di e tri carboxílicos aceleravam a formação de CO2 em diversos tecidos mas não eram consumidos na reação. 2- algum composto de 2 carbonos vindo provavelmente da glicólise se combinava com oxaloacetato e formava um composto de 6 carbonos (citrato) que iniciava uma via de interconversão, Krebs conclui e postula um modelo que ele chamou de “Ciclo do Ácido Cítrico” ou dos “Ácidos Tricarboxílicos”. Krebs então postula que: “O piruvato, ou um derivado vindo da glicólise (acetato), se condensa com o oxaloacetato e forma citrato. Por uma sequência de reações que envolvem cis-aconitato, isocitrato, α-cetoglutarato, succinato, fumarato, malato e oxaloacetato como intermediários, um ácido acético é oxidado e o oxaloacetato necessário para a reação inicial de condensação é regenerado. Isso explica a ação catalítica dos ácidos di e tricarboxílicos (de 4, 5 e 6 carbonos), bem como a capacidade que esses ácidos possuem de se oxidar nos tecidos que oxidam carboidratos.”
 que iniciava uma via de interconversão, Krebs conclui e postula um modelo que ele chamou de Ciclo do Ácido Cítrico ou dos Ácidos Tricarboxílicos . Krebs então postula que: O piruvato, ou um derivado vindo da glicólise (acetato), se condensa com o oxaloacetato e forma citrato. Por uma sequência de reações que envolvem cis-aconitato, isocitrato, α-cetoglutarato, succinato, fumarato, malato e oxaloacetato como intermediários, um ácido acético é oxidado e o oxaloacetato necessário para a reação inicial de condensação é regenerado. Isso explica a ação catalítica dos ácidos di e tricarboxílicos (de 4, 5 e 6 carbonos), bem como a capacidade que esses ácidos possuem de se oxidar nos tecidos que oxidam carboidratos.")
6
Os experimentos de Fritz Lipmann
Naquela época já se sabia que a Coenzima A estava envolvida em reações de transferência de carbono e que ela era derivado do ácido pantotênico, uma vitamina pertencente ao complexo B (vit. B5). - A síntese de citrato aumenta na presença de CoA Ac. Pantotênico = Vit. B5 - Lipmann demonstra que o consumo de O2 é aumentado na presença de CoA quando se fornece glicose-fosfato para leveduras
. - A síntese de citrato aumenta na presença de CoA. Ac. Pantotênico = Vit. B5. - Lipmann demonstra que o consumo de O2 é aumentado na presença de CoA quando se fornece glicose-fosfato para leveduras.")
7
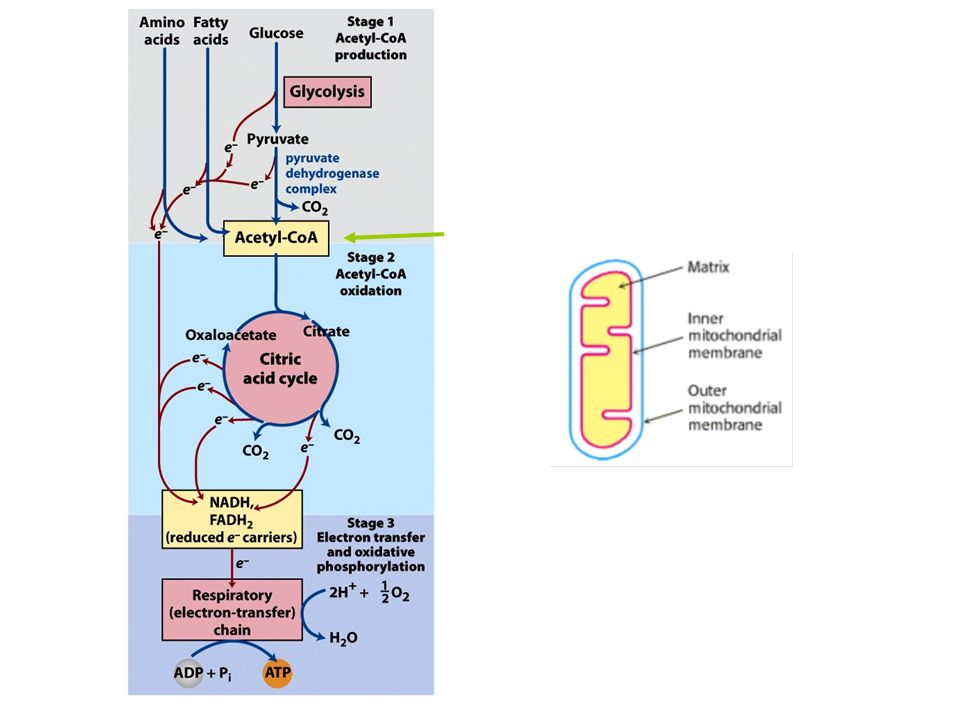
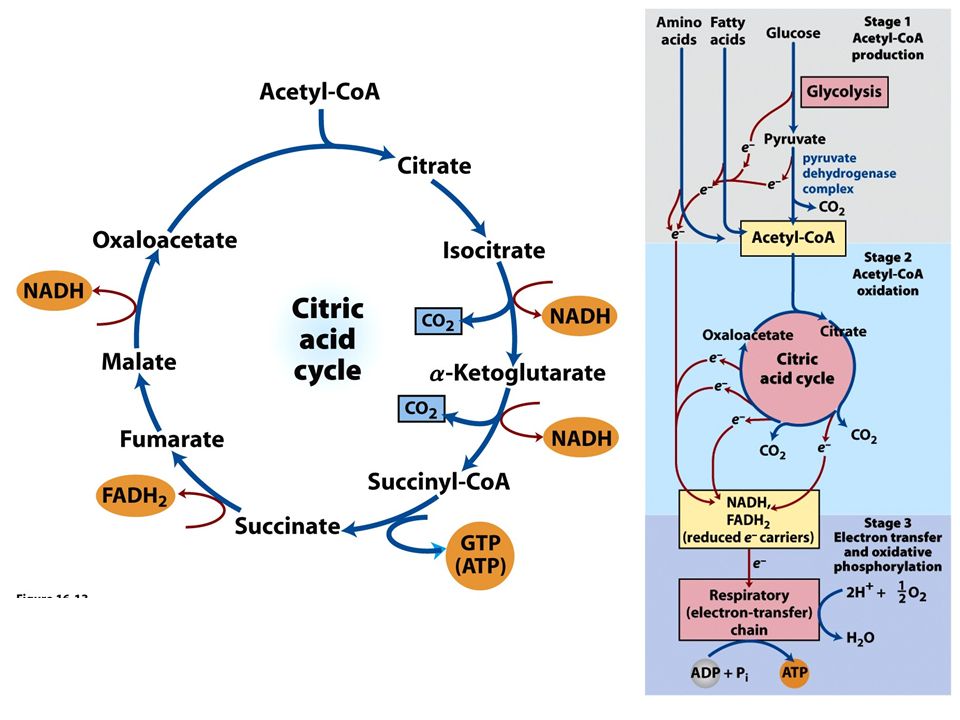
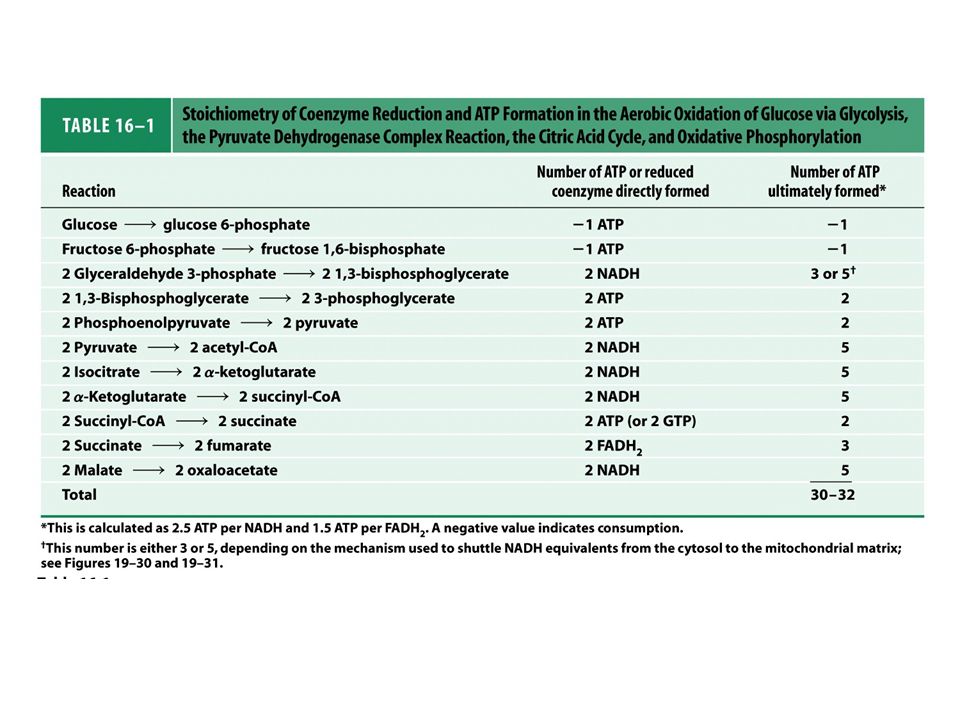
Ciclo do ácido cítrico ou ciclo de krebs
FIGURE 16-1 Catabolism of proteins, fats, and carbohydrates in the three stages of cellular respiration. Stage 1: oxidation of fatty acids, glucose, and some amino acids yields acetyl-CoA. Stage 2: oxidation of acetyl groups in the citric acid cycle includes four steps in which electrons are abstracted. Stage 3: electrons carried by NADH and FADH2 are funneled into a chain of mitochondrial (or, in bacteria, plasma membrane-bound) electron carriers—the respiratory chain—ultimately reducing O2 to H2O.This electron flow drives the production of ATP.
 electron carriers—the respiratory chain—ultimately reducing O2 to H2O.This electron flow drives the production of ATP.")
8
Complexo piruvato desidrogenase:
E1= piruvato desidrogenase E2= dihidrolipoil transacetilase E3= dihidrolipoil desidrogenase

9
FIGURE 16-3 Coenzyme A (CoA)
FIGURE 16-3 Coenzyme A (CoA). A hydroxyl group of pantothenic acid is joined to a modified ADP moiety by a phosphate ester bond, and its carboxyl group is attached to β-mercaptoethylamine in amide linkage. The hydroxyl group at the 3′ position of the ADP moiety has a phosphoryl group not present in free ADP. The —SH group of the mercaptoethylamine moiety forms a thioester with acetate in acetylcoenzyme A (acetyl-CoA) (lower left). FIGURE 16-4 Lipoic acid (lipoate) in amide linkage with a Lys residue. The lipoyllysyl moiety is the prosthetic group of dihydrolipoyl transacetylase (E2 of the PDH complex). The lipoyl group occurs in oxidized (disulfide) and reduced (dithiol) forms and acts as a carrier of both hydrogen and an acetyl (or other acyl) group.
. A hydroxyl group of pantothenic acid is joined to a modified ADP moiety by a phosphate ester bond, and its carboxyl group is attached to β-mercaptoethylamine in amide linkage. The hydroxyl group at the 3′ position of the ADP moiety has a phosphoryl group not present in free ADP. The —SH group of the mercaptoethylamine moiety forms a thioester with acetate in acetylcoenzyme A (acetyl-CoA) (lower left). FIGURE 16-4 Lipoic acid (lipoate) in amide linkage with a Lys residue. The lipoyllysyl moiety is the prosthetic group of dihydrolipoyl transacetylase (E2 of the PDH complex). The lipoyl group occurs in oxidized (disulfide) and reduced (dithiol) forms and acts as a carrier of both hydrogen and an acetyl (or other acyl) group.")
10
Beriberi- falta de vitamina B1
Beriberi- falta de vitamina B1. Animais deficientes em tiamina são incapazes de oxida o piruvato!

11
FIGURE 16-6 Oxidative decarboxylation of pyruvate to acetyl-CoA by the PDH complex. The fate of pyruvate is traced in red. In step 1 pyruvate reacts with the bound thiamine pyrophosphate (TPP) of pyruvate dehydrogenase (E1), undergoing decarboxylation to the hydroxyethyl derivative (see Figure 14-14). Pyruvate dehydrogenase also carries out step 2, the transfer of two electrons and the acetyl group from TPP to the oxidized form of the lipoyllysyl group of the core enzyme, dihydrolipoyl transacetylase (E2), to form the acetyl thioester of the reduced lipoyl group. Step 3 is a transesterification in which the ムSH group of CoA replaces the —SH group of E2 to yield acetyl-CoA and the fully reduced (dithiol) form of the lipoyl group. In step 4 dihydrolipoyl dehydrogenase (E3) promotes transfer of two hydrogen atoms from the reduced lipoyl groups of E2 to the FAD prosthetic group of E3, restoring the oxidized form of the lipoyllysyl group of E2. In step 5 the reduced FADH2 of E3 transfers a hydride ion to NAD+, forming NADH. The enzyme complex is now ready for another catalytic cycle. (Subunit colors correspond to those in Figure 16-5b.)
 of pyruvate dehydrogenase (E1), undergoing decarboxylation to the hydroxyethyl derivative (see Figure 14-14). Pyruvate dehydrogenase also carries out step 2, the transfer of two electrons and the acetyl group from TPP to the oxidized form of the lipoyllysyl group of the core enzyme, dihydrolipoyl transacetylase (E2), to form the acetyl thioester of the reduced lipoyl group. Step 3 is a transesterification in which the ムSH group of CoA replaces the —SH group of E2 to yield acetyl-CoA and the fully reduced (dithiol) form of the lipoyl group. In step 4 dihydrolipoyl dehydrogenase (E3) promotes transfer of two hydrogen atoms from the reduced lipoyl groups of E2 to the FAD prosthetic group of E3, restoring the oxidized form of the lipoyllysyl group of E2. In step 5 the reduced FADH2 of E3 transfers a hydride ion to NAD+, forming NADH. The enzyme complex is now ready for another catalytic cycle. (Subunit colors correspond to those in Figure 16-5b.).")
12
The functional groups are screened in the structural formula
The functional groups are screened in the structural formula. In the space-filling model, N is blue, C is black, P is orange, O is red, and H is white. The yellow atom at the left is the sulfur of the critical thioester bond between the acetyl moiety and coenzyme A

13
Inibição pelo produtos: NADH e acetil-CoA.
Estes ativam piruvato desidrogenase cinase (mamífero) => fosforila E1 => inativa ATP ativa piruvato desidrogenase cinase Piruvato e ADP inibem a piruvato desidrogenase cinase Ca2+ inibe piruvato desidrogenase cinase e ativa piruvato desidrogenase fosfatase. Insulina => ativa a piruvato desidrogenase fosfatase => ativa E1
 => fosforila E1 => inativa. ATP ativa piruvato desidrogenase cinase. Piruvato e ADP inibem a piruvato desidrogenase cinase. Ca2+ inibe piruvato desidrogenase cinase e ativa piruvato desidrogenase fosfatase. Insulina => ativa a piruvato desidrogenase fosfatase => ativa E1.")
15
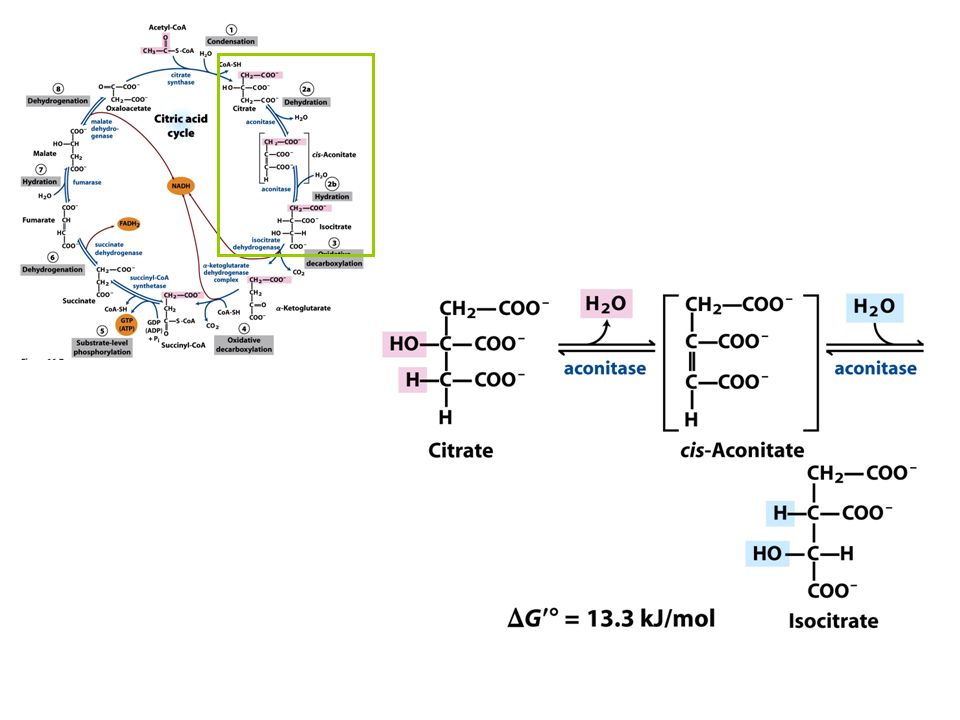
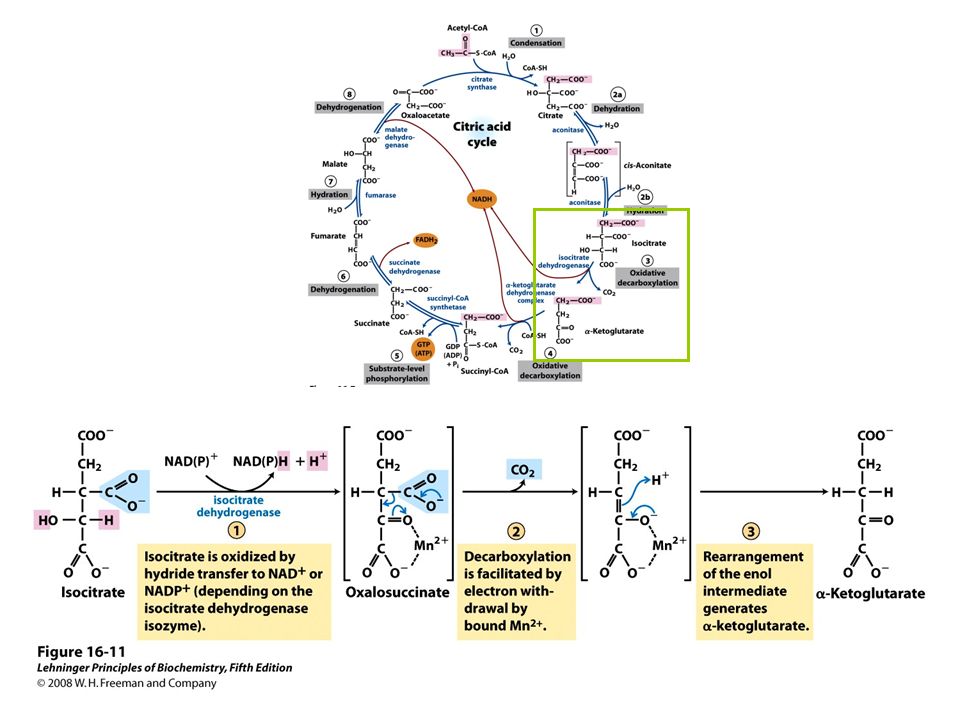
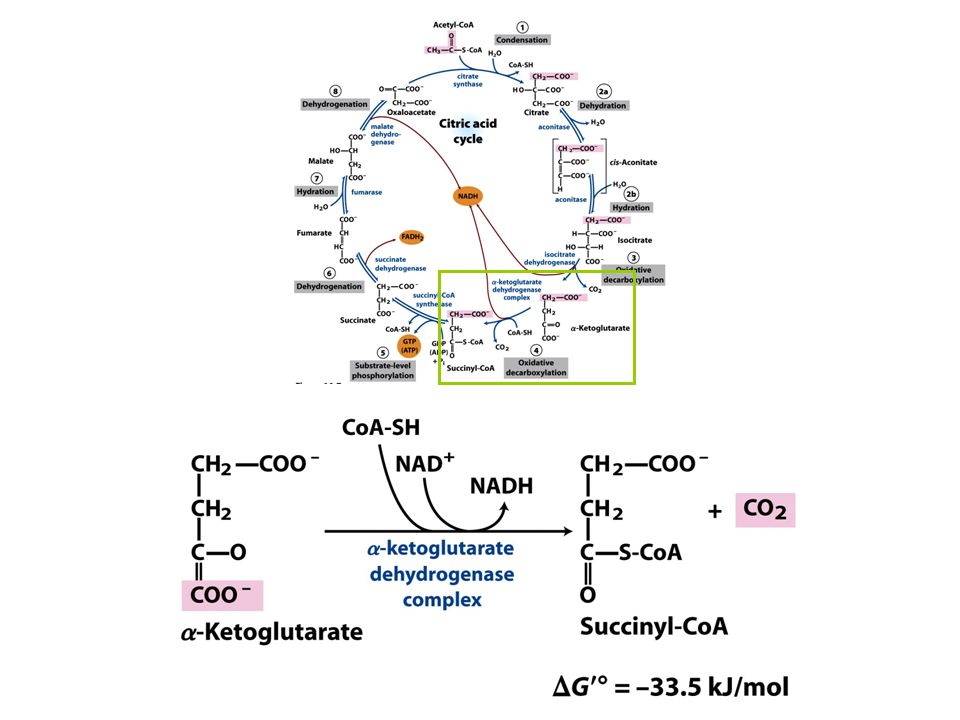
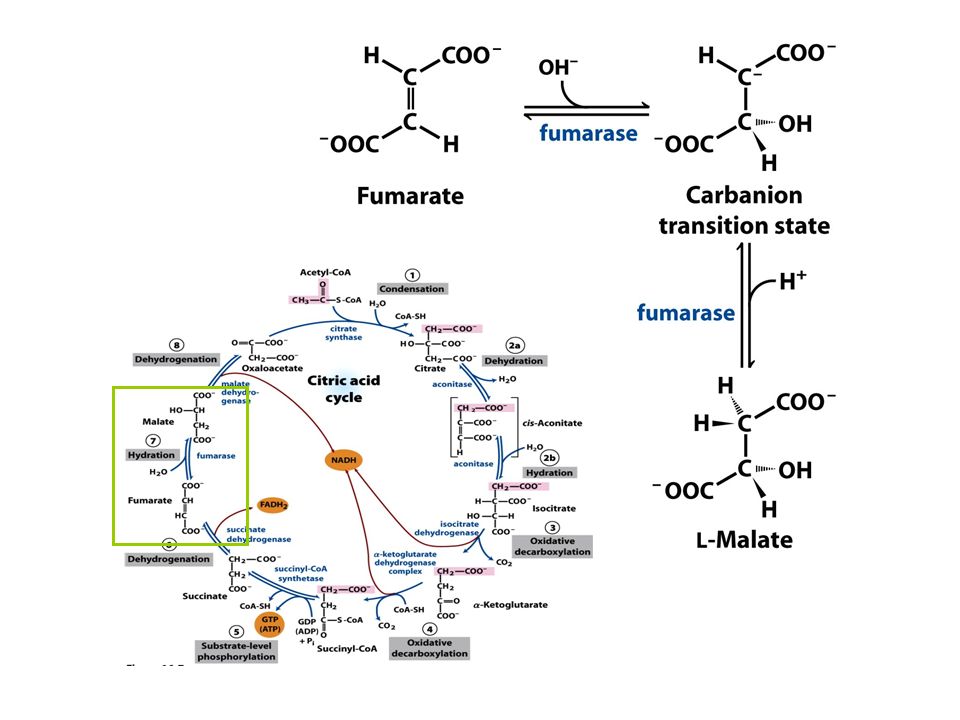
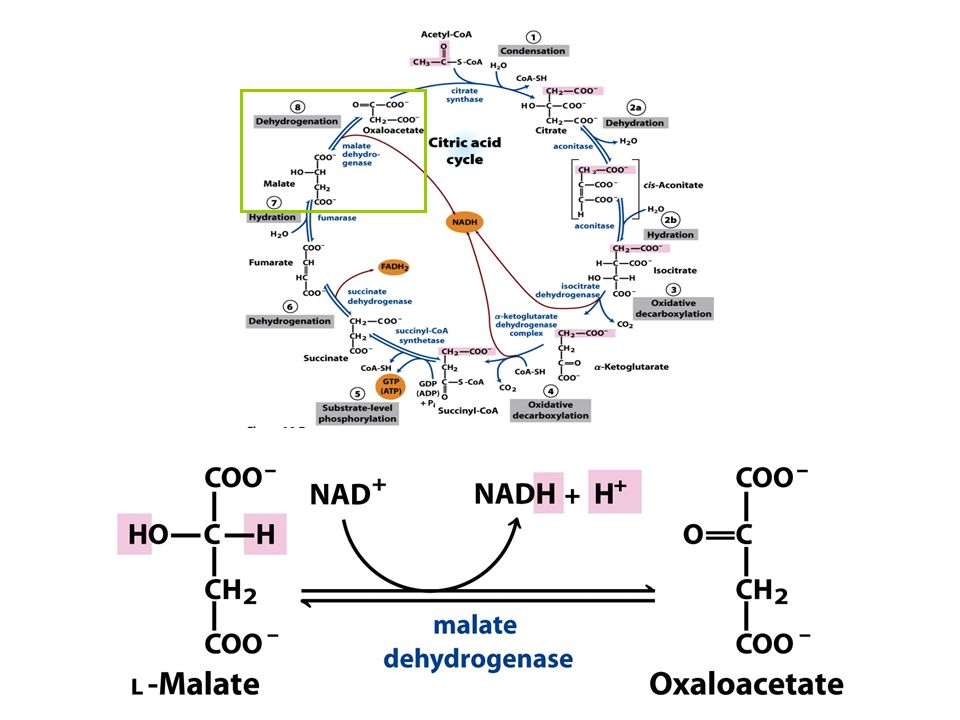
FIGURE 16-7 Reactions of the citric acid cycle
FIGURE 16-7 Reactions of the citric acid cycle. The carbon atoms shaded in pink are those derived from the acetate of acetyl-CoA in the first turn of the cycle; these are not the carbons released as CO2 in the first turn. Note that in succinate and fumarate, the two-carbon group derived from acetate can no longer be specifically denoted; because succinate and fumarate are symmetric molecules, C-1 and C-2 are indistinguishable from C-4 and C-3. The number beside each reaction step corresponds to a numbered heading on pages 622–628. The red arrows show where energy is conserved by electron transfer to FAD or NAD+, forming FADH2 or NADH + H+. Steps 1, 3, and 4 are essentially irreversible in the cell; all other steps are reversible. The product of step 5 may be either ATP or GTP, depending on which succinyl-CoA synthetase isozyme is the catalyst.

16
Sintase: reação de condensação sem nucleosídeo trifosfato (ATP, GTP...)
")
17
Importância da ligação tioester
FIGURE Hydrolysis of acetyl-coenzyme A. Acetyl-CoA is a thioester with a large, negative, standard free energy of hydrolysis. Thioesters contain a sulfur atom in the position occupied by an oxygen atom in oxygen esters. The complete structure of coenzyme A (CoA, or CoASH) is shown in Figure FIGURE Free energy of hydrolysis for thioesters and oxygen esters. The products of both types of hydrolysis reaction have about the same free-energy content (G), but the thioester has a higher free-energy content than the oxygen ester. Orbital overlap between the O and C atoms allows resonance stabilization in oxygen esters; orbital overlap between S and C atoms is poorer and provides little resonance stabilization.
 is shown in Figure FIGURE Free energy of hydrolysis for thioesters and oxygen esters. The products of both types of hydrolysis reaction have about the same free-energy content (G), but the thioester has a higher free-energy content than the oxygen ester. Orbital overlap between the O and C atoms allows resonance stabilization in oxygen esters; orbital overlap between S and C atoms is poorer and provides little resonance stabilization.")
22
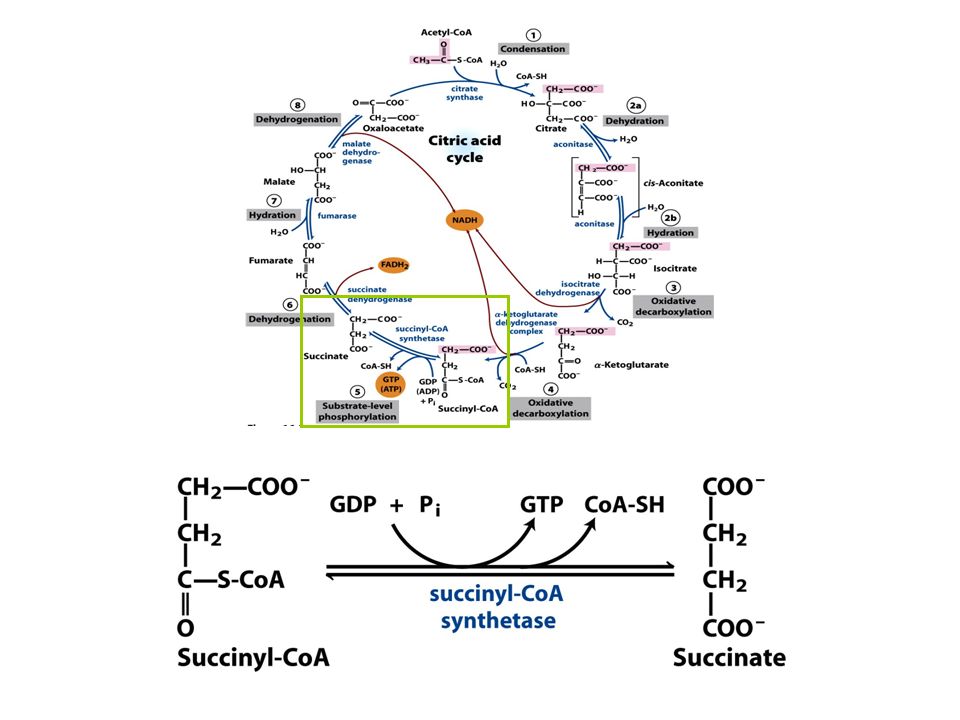
Sintetase: reação de condesação com uso de nucleosídeos trifosfato
FIGURE 16-12a The succinyl-CoA synthetase reaction. (a) In step 1 a phosphoryl group replaces the CoA of succinyl-CoA bound to the enzyme, forming a high-energy acyl phosphate. In step 2 the succinyl phosphate donates its phosphoryl group to a His residue of the enzyme, forming a high-energy phosphohistidyl enzyme. In step 3 the phosphoryl group is transferred from the His residue to the terminal phosphate of GDP (or ADP), forming GTP (or ATP).
 In step 1 a phosphoryl group replaces the CoA of succinyl-CoA bound to the enzyme, forming a high-energy acyl phosphate. In step 2 the succinyl phosphate donates its phosphoryl group to a His residue of the enzyme, forming a high-energy phosphohistidyl enzyme. In step 3 the phosphoryl group is transferred from the His residue to the terminal phosphate of GDP (or ADP), forming GTP (or ATP).")
23
Malonato= inibidor competitivo

26
Intervalo de 10 minutos

29
O catabolismo de proteínas gera diversos intermediários do ciclo de Krebs

30
Vias catabólicas: convergentes
Vias anabólicas: divergentes Algumas vias são cíclicas, ou seja, um precursor da via é regenerado por meio de uma série de reações.

31
Papel do ciclo do ácido cítrico no anabolismo
FIGURE Role of the citric acid cycle in anabolism. Intermediates of the citric acid cycle are drawn off as precursors in many biosynthetic pathways. Shown in red are four anaplerotic reactions that replenish depleted cycle intermediates (see Table 16-2)
")
32
Quando o ciclo está deficiente em oxalacetato e outros intermediários

33
A glicólise e ciclo de krebs estão integrados.
FIGURE Regulation of metabolite flow from the PDH complex through the citric acid cycle in mammals. The PDH complex is allosterically inhibited when [ATP]/[ADP], [NADH]/[NAD+], and [acetyl-CoA]/[CoA] ratios are high, indicating an energy-sufficient metabolic state. When these ratios decrease, allosteric activation of pyruvate oxidation results. The rate of flow through the citric acid cycle can be limited by the availability of the citrate synthase substrates, oxaloacetate and acetyl-CoA, or of NAD+, which is depleted by its conversion to NADH, slowing the three NAD-dependent oxidation steps. Feedback inhibition by succinyl-CoA, citrate, and ATP also slows the cycle by inhibiting early steps. In muscle tissue, Ca2+ signals contraction and, as shown here, stimulates energy-yielding metabolism to replace the ATP consumed by contraction.

34
Diabetes Mellitus: comum no Brasil (prevalência 7,6 % da população brasileira entre 30 e 69 anos)
Apresentam hiperglicemia Tipo I: insulino-dependente ou juvenil. É uma doença auto-imune que provoca a destruição de células b das ilhotas do pâncreas. Tipo II: não insulino-dependente (resistente a insulina, e por secreção deficiente de insulina). 80% estão acima do peso adequado Síndrome Metabólica No diabetes o organismo comporta-se como no jejum prolongado. Um dos métodos de monitoramento da hiperglicemia é o exame que mede a Hemoglobina glicada (HbA1c): em diabetes essa taxa pode ser até 3 X maior.
. 80% estão acima do peso adequado Síndrome Metabólica. No diabetes o organismo comporta-se como no jejum prolongado. Um dos métodos de monitoramento da hiperglicemia é o exame que mede a Hemoglobina glicada (HbA1c): em diabetes essa taxa pode ser até 3 X maior.")
35
FIGURE 17-18 Formation of ketone bodies from acetyl-CoA
FIGURE Formation of ketone bodies from acetyl-CoA. Healthy, well-nourished individuals produce ketone bodies at a relatively low rate. When acetyl-CoA accumulates (as in starvation or untreated diabetes, for example), thiolase catalyzes the condensation of two acetyl-CoA molecules to acetoacetyl-CoA, the parent compound of the three ketone bodies. The reactions of ketone body formation occur in the matrix of liver mitochondria. The six-carbon compound β-hydroxy-β-methylglutaryl-CoA (HMG-CoA) is also an intermediate of sterol biosynthesis, but the enzyme that forms HMG-CoA in that pathway is cytosolic. HMG-CoA lyase is present only in the mitochondrial matrix. FIGURE Ketone body formation and export from the liver. Conditions that promote gluconeogenesis (untreated diabetes, severely reduced food intake) slow the citric acid cycle (by drawing off oxaloacetate) and enhance the conversion of acetyl-CoA to acetoacetate. The released coenzyme A allows continued β oxidation of fatty acids.
, thiolase catalyzes the condensation of two acetyl-CoA molecules to acetoacetyl-CoA, the parent compound of the three ketone bodies. The reactions of ketone body formation occur in the matrix of liver mitochondria. The six-carbon compound β-hydroxy-β-methylglutaryl-CoA (HMG-CoA) is also an intermediate of sterol biosynthesis, but the enzyme that forms HMG-CoA in that pathway is cytosolic. HMG-CoA lyase is present only in the mitochondrial matrix. FIGURE Ketone body formation and export from the liver. Conditions that promote gluconeogenesis (untreated diabetes, severely reduced food intake) slow the citric acid cycle (by drawing off oxaloacetate) and enhance the conversion of acetyl-CoA to acetoacetate. The released coenzyme A allows continued β oxidation of fatty acids.")
36
Prática I - Correção

Apresentações semelhantes






 ou ANAeróbia (outras substâncias);>")












