Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Transporte vesicular: transporte de substâncias através de vesículas Aparelho de Golgi

2
Transporte vesicular Ptns recém sintetizadas entram na rota biossintética secretora do RE; Ptnas passam por compartimentos onde são modificadas; Vesículas que transportam uma carga determinada saem de um compartimento doador e se fusionam a um compartimento aceptor;

3
Aparelho de Golgi É um dos principais sítios de síntese de carboidratos, bem como, uma estação de classificação e de destinação para produtos do RE; Grande proporção dos carboidratos que são produzidos no golgi são conectados como cadeias laterais de oligossacarídeos em muitas ptns e lipídeos que o RE envia;

4
O lúmen de cada compartimento é topologicamente equivalente ao exterior da célula e esses compartimentos estão todos em constante comunicação;

5
O complexo de Golgi é uma organela com membrana, formada por várias compartimentos ordenados na forma de pratos empilhados; Cada pilha de Golgi é composta de 2 faces distintas: uma de entrada (Cis) e outra de saída (Trans). Ambas estão conectadas a compartimentos especiais (Cis-Golgi-Network e Trans-Golgi-Network), que são compostos por uma rede de estruturas tubulares em forma de cisterna
 e outra de saída (Trans). Ambas estão conectadas a compartimentos especiais (Cis-Golgi-Network e Trans-Golgi-Network), que são compostos por uma rede de estruturas tubulares em forma de cisterna.")
6
1 - Porção cis Golgi Network= voltada para o RE rugoso = recebe vesículas;
2 – Porção média = entre cis/trans = só processamento; 3 – Porção trans Golgi Network = voltada para a membrana plasmática = envia vesículas. A comunicação entre compartimentos é por meio de vesículas de transferência.

7
É no Golgi que ocorre a síntese de carboidratos e é a estação de separação e distribuição dos produtos do RE; Muitos carboidratos celulares são sintetizados no Golgi: Pectina e Hemicelulose da parede celular de células vegetais e as glicosaminoglicanas na matriz extracelular das células animais; Uma grande proporção dos carboidratos que o Golgi produz estão ligados a oligossacarídeos de proteínas e lipídios provenientes do RE; Certos oligossacarídeos servem como marcas para o direcionamento específico de proteínas para diferentes destinos.

8
O Golgi é abundante em células que são especializadas em secreção
O Golgi é abundante em células que são especializadas em secreção. Exemplo: as células caliciformes (globet) do intestino, que produzem grande quantidade de muco rico em polissacarídeo na luz do intestino. Nessas células, grandes vesículas são encontradas na parte do Trans-Golgi-Network.
 do intestino, que produzem grande quantidade de muco rico em polissacarídeo na luz do intestino. Nessas células, grandes vesículas são encontradas na parte do Trans-Golgi-Network.")
9
O transporte de vesículas do RE para o Golgi se dá a partir de vesículas, nos chamados elementos de transição do RE, que são quase totalmente livres de ribossomos e que fundem-se à membrana do Cis-Golgi-Network, transferindo proteínas e lipídios sintetizadas pelo RE.

10
O tráfego entre RE e o Golgi é bidirecional
O tráfego de retorno tem duas funções principais: Manter quantidades estáveis de membranas em cada compartimento Recuperar proteínas do compartimento doador que são necessárias para o seu normal funcionamento

11
Proteínas residentes do RE são seletivamernte recolocadas ou redirecionadas para o RE a partir do Cis-Golgi-Network. KDEL (Lisina, ácido aspártico, ácido Gluâmico e Leucina) é o sinal de retenção no RE.
 é o sinal de retenção no RE..")
12
Tratada com Brefeldina A
Proteínas do Golgi retornam para o RE quando a célula é tratada com a droga Brefeldina A que promove a ruptura do Golgi. Tratada com Brefeldina A Normal Manosidase

13
A saída do RE pode ser considerada como ponto de controle de qualidade de proteínas. Se a conformação de uma proteína ou montagem das sub-unidades protéicas não for completada no RE, a proteína é degradada.

15
Duas classes de oligossacarídeos N-ligados estão associadas a glicoproteínas de mamíferos: Oligossacarídeos ricos em manose e oligossacarídeos complexos;

16
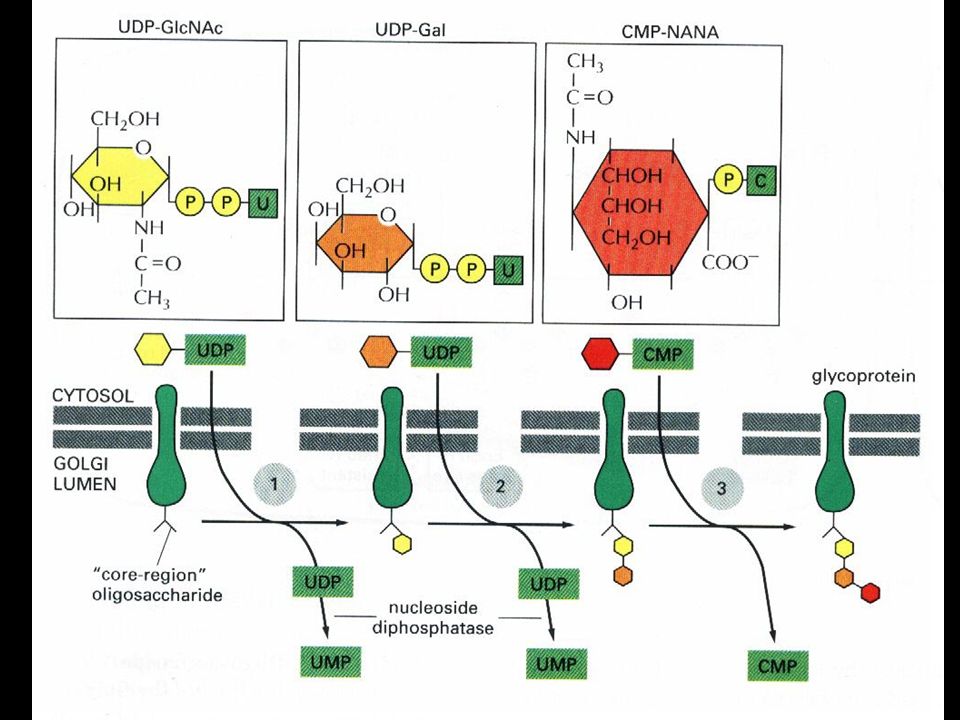
As cadeias de oligossacarídeos são processadas no Golgi

18
As cisternas do Golgi são organizadas como uma série de compartimentos de processamento:
Cis= contínuo com o Cis-Golgi-Network; Medial= cisterna central; Trans= contínuo com o Trans-Golgi-Network.

19
A diferença funcional entre as subdivisões do Golgi Cis, Medial e Trans foi a descoberta, através da localização, em regiões distintas, de enzimas responsáveis pelo processamento dos oligossacarídeos N-ligados. Corado com ósmio Não corado Identificação de enzimas

20
Muitas proteínas são modificadas pela adição de oligossacarídeos em grupos OH de cadeias laterais de serina e treonina: Esses oligossacarídeos são denominadaos de O-ligados; Os carboidratos da membrana celular estão voltados para o lado da membrana que é topologicamente equivalente ao lado de fora da célula.

21
Qual a função da glicosilação?

22
Maioria das funções ainda são desconhecidas;
Tornar uma glicoproteína mais resistente a ação de proteases; Direcionar o transporte de ptns; Regulação;

23
Lisossomos

24
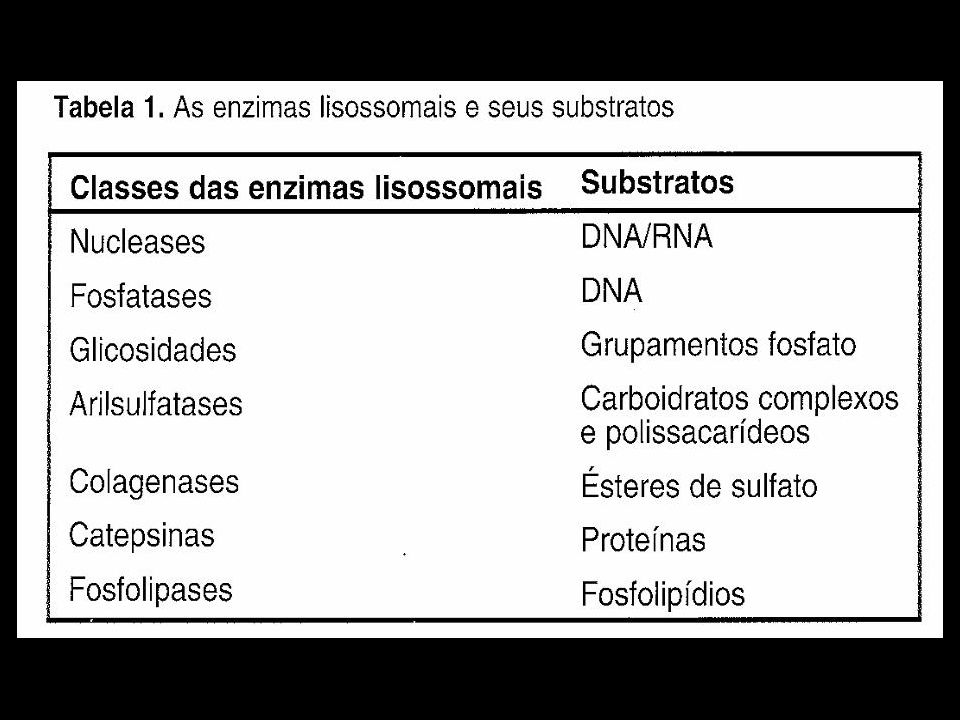
Lisossomos São organelas citoplasmáticas, onde ocorre a maioria da digestão intracelular; Existem mais de 40 tipos de enzimas hidrolíticas ácidas (pH=5.0); A membrana do lisossomo é rica em glicoproteínas e proteínas de transporte dos produtos de degradação.
; A membrana do lisossomo é rica em glicoproteínas e proteínas de transporte dos produtos de degradação.")
27
digestão intracelular.
Função: digestão intracelular. Quebra de restos intra e extracelulares, destruição de microorganismos fagocitados e produção de nutrientes para célula O que é digerido???? Material de endocitose; Organelas e moléculas (reciclagem).
.")
28
Estrutura Podem acumular material não digerido corpo residual.
Esféricas e de tamanho variável. Formadas por uma bicamada lipídica. Identificação = fosfatase ácida. Podem acumular material não digerido corpo residual. Face interna revestimento de carboidratos proteção contra a ação do conteúdo lisossomal.

29
Formação dos lisossomos
A partir do complexo de Golgi. 1 – A partir da face trans vesículas com enzimas pré-lisossomais. 2 - Endossomos formação de pH ácido bombas de prótons (próton-ATPases): Resultados: pH abaixo de 6; Dissociação das pré-enzimas dos receptores.
: Resultados: pH abaixo de 6; Dissociação das pré-enzimas dos receptores.")
30
Lisossomos

31
pH de lisossomos e endossomos
Visualização histoquímica dos lisossomos. Precipitado de fosfato de chumbo para localizar a fostatase ácida. pH de lisossomos e endossomos V= pH 5.0 Lis., A e Ve= pH liso e endo 6.5. Proteínas são marcadas com subst. Fluorescente e fagocitadas.

32
Vacúolos de plantas e fungos são lisossomos versáteis; de 30 a 90% do vol. Celular;
Várias funções: Estoque de nutrientes; produtos de excreção; compartimento de degradação.

33
Materiais são transportados para os lisossomos por diversos caminhos;
Três caminhos de degradação encontram-se nos lisossomos: 1. Endocitose; 2. Autofagia e 3. Fagocitose; Os lisossomos são centrais de encontro.

34
Autofagia Uma mitocôndria possui meia vida de 10 dias no fígado;
O processo de autofagia começa com o empacotamento da organela por membranas de RE criando um autofagossomo que então se funde com os lisossomos;

35
Como as proteínas lisossômicas são reconhecidas no Aparelho de Golgi?

36
Enzimas lisossomais são separadas de outras proteínas no Trans-Golgi-Network por proteína receptora ligada à membrana que reconhece manose-6-fosfato (que foi introduzida no cis-Golgi-Network). Receptores de M-6-F na membrana do Trans-Golgi-Network formam vesículas especiais de transporte cobertas por clatrina; O receptor se liga à M-6-P em pH=7 no Trans-Golgi-Network e se solta a pH= 6 no endossomo tardio.

37
Síntese do marcador de manose-6-fosfato na hidrolase lisossomal

38
Um sinal derivado da estrutura tridimensional da proteína é necessário para o reconhecimento pela N-acetil-glicosamina fosfotransferase; Defeitos na enzima N-acetil-glicosamina fosfotransferase causa doenças de depósito;

39
Síntese das enzimas lisossomais
1 - Síntese a partir dos ribossomos formação da pré-enzima; 2 – No retículo endoplasmático glicosilação da pré-enzima; Adição de açúcar no resíduo de asparagina (N-ligado). 3 – Complexo de Golgi Na rede cis adição de manose-6-P na pré-enzima. Na rede trans ligação aos receptores. 4 – Endossomos pH ácido desligamento dos receptores. Ativação das enzimas.
. 3 – Complexo de Golgi. Na rede cis adição de manose-6-P na pré-enzima. Na rede trans ligação aos receptores. 4 – Endossomos pH ácido desligamento dos receptores. Ativação das enzimas.")
40
depois da liberação das enzimas os receptores retornam para rede Trans do golgi e parecem ter um peptídeo sinal KDEL;

41
Endocitose

42
Fagocitose ingestão de microorganismos ou debris celulares via fagossomos > ou = a 250 nm em diâmetro; Fagocitose de hemáceas alteradas por um macrófago.

43
Células fagocíticas especias denominadas de fagócitos profissionais (macrófagos e neutrófilos) podem ingerir partículas grandes. Os macrófagos combatem infecção e removem células danificadas (bilhões de hemácias velhas são removidas por dia) Um macrófago ingere 25% de seu volume em 1 h; Nos protozoários a fagocitose é uma forma de alimentação;
 Um macrófago ingere 25% de seu volume em 1 h; Nos protozoários a fagocitose é uma forma de alimentação;")
44
Pinocitose Ingestão de fluidos e solutos via pequenas vesículas < ou = a 150 nm em diâmetro; Cél. Realiza pinocitose constantemente; Ciclo endocítico-exocítico; As vesículas pinocíticas se formam a partir de fossas cobertos por uma proteína denominada CLATRINA na membrana plasmática;

45
Estrutura de uma capa de clatrina
Formados por cadeias polipeptídicas: 3 grandes e 3 pequenas Triquélions

46
Formação das vesículas cobertas por clatrina

47
Peptídeo-sinal para endocitose

48
Formação de vesículas de cobertas por clatrina

49
Por volta de 2500 vesículas coberta por clatrina deixam a MP de fibroblastos em cultura a cada minuto; A endocitose poder ser mediada por receptor (ex: absorção de colesterol); A maioria do colesterol no sangue é ligado a proteína na forma de partículas conhecidas como "Low-Density-Lipoproteins" ou LDL. Quando a célula necessita de colesterol o receptor é sintetizado e enviado à MP. Mais de 25 tipos diferentes de receptores participam da endocitose mediada por receptor.
; A maioria do colesterol no sangue é ligado a proteína na forma de partículas conhecidas como Low-Density-Lipoproteins ou LDL. Quando a célula necessita de colesterol o receptor é sintetizado e enviado à MP. Mais de 25 tipos diferentes de receptores participam da endocitose mediada por receptor.")
50
defeitos na entrada de LDL provocam arteriosclerose

51
Revestimento Vesicular
Exisem 3 tipos de vesículas cobertas bem caracterizadas, que diferem na capa protéica que possuem: vesículas cobertas por clatrina, cobertas por COPI-e cobertas por COPII cobertas por clatrina (Transporte seletivo de receptores transmembrana, receptor M6P na memb do TransGolgi Network, receptor de LDL da MP);
;")
52
Proteínas que revestem vesículas

53
Transporte seletivo por vesículas cobertas por clatrina
Transporte seletivo por vesículas cobertas por clatrina. As proteínas adaptinas ligam-se à receptores de carga e às clatrinas

54
Dinamina

55
2. Vesículas cobertas por COPI- e COPII medeiam, comumente, o transporte do RE e Golgi

56
Modelo da formação da vesícula coberta com coatomer (COPII)
Ação da brefeldina A

57
GTPases Controlam a montagem da capa protéica das vesículas
Ação da brefeldina A

58
Partículas endocitadas ligadas à receptores podem ter destinos diferentes:
1 – reciclados e devolvidos para o domínio da MP de onde vieram; 2 – seguir para um domínio diferente da MP mediado por um processo chamado de TRANSCITOSE; 3 – seguir para o lisossomo e serem degradados

59
As proteínas receptoras podem ter 3 destinos diferentes na endocitose:
1) retornam p/ MP ex. Receptor de LDL; Transferina 2) lisossomos; 3) novo domínio na MP (transcitose) ex. EGF (fator de crescimento epidermal
 retornam p/ MP ex. Receptor de LDL; Transferina. 2) lisossomos; 3) novo domínio na MP (transcitose) ex. EGF (fator de crescimento epidermal.")
60
Caminho endocítico da MP para o lisossomo;
Células epiteliais possuem endossomos iniciais e tardios;

61
Dois compartimentos endossomais iniciais em células epiteliais

62
Exocitose

64
Secreção constitutiva e regulada

65
Três classes de ptnas são separadas no Golgi:
Destinadas aos lisossomos: são marcadas com M6P; Proteínas com secreção regulada: marcadas por sinais análogos ao de MP; Maioria das outras ptnas: são transportadas para a superfície celular pela rota secretora constitutiva não-seletiva (rota padrão);
;")
66
O local mais conhecido de direcionamento de substâncias dentro da célula é na rede Trans do Golgi;

67
Partícula de Ac-ouro ligada à clatrina (vesícula imatura com pro-insulina)
Formação de vesículas secretoras brotando do trans-golgi-network (células b do pâncreas) Vesícula secretora madura (sem ouro) Exocitose de vesículas secretoras Insulina
 Vesícula secretora madura (sem ouro) Exocitose de vesículas secretoras. Insulina.")
68
Proteínas são, muitas vezes, processadas proteolíticamente durante a formação de vesículas secretoras. ex. pro-hormônio propiocortina secretado pela glândula pituitária (hipófise); Vesículas secretoras esperam perto das MP até serem sinalizadas a liberar seus conteúdos. Produzido:cél. lobo anterior Cél. Lobo intern. (pituitária)
; Vesículas secretoras esperam perto das MP até serem sinalizadas a liberar seus conteúdos. Produzido:cél. lobo anterior. Cél. Lobo intern. (pituitária)")
70
Mastócito de rato com vesículas de histamina.
A exocitose regulada é uma resposta local da MP é no citoplasma adjacente (um sinal para secreção é, na maioria das vezes,um mensageiro químico (p.ex., um hormônio que se liga à membrana plasmática); No caso do axônio, é um sinal elétrico que abre canais iônicos de Ca2+. Os componentes da membrana de vesículas secretoras são reciclados. Mastócito de rato com vesículas de histamina. Esfera
; No caso do axônio, é um sinal elétrico que abre canais iônicos de Ca2+. Os componentes da membrana de vesículas secretoras são reciclados. Mastócito de rato com vesículas de histamina. Esfera.")
71
Vesículas sinápticas se formam a partir de endossomos
Vesículas sinápticas se formam a partir de endossomos. Muitos neurônios “disparam” mais de 1000x por segundo, liberando vesículas sinápticas toda vez.

72
Células polarizadas direcionam proteínas do rede Trans do golgi para o domínio apropriado na membrana (Transcitose); Junções compactas

73
Mecanismos moleculares do transporte vesicular e manutenção da diversidade de compartimentos celulares.

74
Como a célula mantém a diversidade dos compartimentos com as trocas massivas de substâncias entre eles? por marcadores nas membranas

75
Papel postulado das proteínas SNAREs na condução do transporte vesicular ( adaptadores de fusão vesicular) Proteínas SNARE no SN são alvos das toxinas do botulismo e tétano.

76
Dissociação de SNARE

77
Papel postulado das proteínas Rab conferindo especificidade no ancoramento da vesícula de transporte: - fixação e ancoragem das vesículas à membrana alvo;

78
Modelo para o mecanismo de fusão de vesículas mediado por proteínas.

79
Entrada do vírus HIV em linfócitos

80
Modelo para a fusão de membranas mediado por uma proteína viral

81
Fusão de células de inseto infectadas por um baculovírus recombinante expressando o gene do envelope do vírus da Febre Amarela A B

Apresentações semelhantes














 e o envoltório nuclear. Formado por um líquido gelatinoso,>")






