Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Fundamentos da expressão gênica em células vegetais Transdução de sinais em plantas Controle do ciclo celular

2
INTRODUÇÃO Crescimento e desenvolvimento de uma planta iniciam com germinação de uma semente. Continuam com desenvolvimento vegetativo, floração, frutificação, dispersão de sementes e processos de senescência. Fase vegetativa do desenvolvimento tem início com célula única, o zigoto, predestinado a crescer de forma organizada. O zigoto passa a sofrer três processos celulares imprescindíveis para o crescimento e o desenvolvimento: divisão celular, alongamento celular e diferenciação celular. Estes processos produzem a organogênese. Nas plantas, a organogênese se mantém constante nos meristemas, o que é conhecido como crescimento indeterminado. Crescimento e desenvolvimento dependem de sinais ambientais (abióticos ou exógenos), principalmente presença de água, nutrição, luz, fotoperíodo e temperaturas. Estes induzem a síntese de sinais endógenos, os hormônios vegetais ou mensageiros primários.
, principalmente presença de água, nutrição, luz, fotoperíodo e temperaturas. Estes induzem a síntese de sinais endógenos, os hormônios vegetais ou mensageiros primários.")
3
Hormônios vegetais ou fitormônios atuam no controle do ciclo celular, da expressão gênica e da ativação enzimática. Os primeiros hormônios vegetais a serem estudados foram: AUXINAS, GIBERELINAS, CITOCININAS, ETILENO E ÁCIDO ABSCÍSICO. Mais tarde foram descobertos: BRASSINOSTERÓIDES, JASMONATOS, ÁCIDO SALICÍLICO, POLIAMINAS, SISTEMINAS E OLIGOSSACARINAS. Ativação” e “repressão” de genes causam alteração nos padrões de síntese de proteínas e enzimas. Estas estão envolvidas na indução de divisão e diferenciação celular.

4
Arabidopsis thaliana (L.) Heynh. (Brassicaceae)
Genes são seqüências de DNA que codificam mRNAs. Estes codificam proteínas estruturais e enzimas. Genoma de plantas contém entre 20 a 30 mil genes. O genoma de Arabidopsis thaliana está totalmente mapeado e contém genes. 1500 codificam proteínas fatores de transcrição responsáveis pela transcrição do gene em seu mRNA. 1200 codificam proteínas quinases (ou cinases), importantes na transdução de sinais biológicos. 600 codificam proteínas quinases receptoras celulares presentes nas membranas. Arabidopsis thaliana (L.) Heynh. (Brassicaceae)
, importantes na transdução de sinais biológicos. 600 codificam proteínas quinases receptoras celulares presentes nas membranas. Arabidopsis thaliana (L.) Heynh. (Brassicaceae)")
5
Esquema de expressão gênica
TRANSCRIÇÃO TRADUÇÃO

6
Genoma de arroz é 5 vezes maior que o de Arabidopsis thaliana, o menor genoma encontrado em plantas até agora. Genes podem ser agrupados em dois grandes grupos: constitutivos e regulados. Genes constitutivos (ou“housekeeping”) são responsáveis pela manutenção de processos celulares vitais, por exemplo, a respiração celular, estando permanentemente ligados. Genes regulados são ativados em fases especiais do desenvolvimento em resposta aos estímulos específicos como luz, fotoperíodo stress. Como exemplo, temos os genes envolvidos com expressão de órgãos florais e genes envolvidos com o aparato fotossintético.
 são responsáveis pela manutenção de processos celulares vitais, por exemplo, a respiração celular, estando permanentemente ligados. Genes regulados são ativados em fases especiais do desenvolvimento em resposta aos estímulos específicos como luz, fotoperíodo stress. Como exemplo, temos os genes envolvidos com expressão de órgãos florais e genes envolvidos com o aparato fotossintético.")
7
Fundamentos da expressão gênica
Expressão de genes eucarióticos requer proteínas especiais, as RNA polimerases. Há três tipos de RNA polimerases: I, II e III. RNA polimerases I - no nucléolo - sintetizam rRNAs. RNA polimerases II - no nucleoplasma, sintetizam pré-mRNA. RNA polimerases III - no nucleoplasma, sintetizam pequenos RNAs: tRNA e 5S rRNA. RNA polimerase II requer para seu funcionamento, proteínas conhecidas como fatores gerais de transcrição que formam o complexo inicial de transcrição.

8
Transcrição inicia quando o último fator geral de transcrição, liga-se ao complexo fosforilando a RNA polimerase II. RNA polimerase II copia apenas uma das fitas de DNA no sentido anti-senso 3’ para 5’. Genes possuem também seqüências adjacentes de DNA: promotor ou promotor mínimo (seqüência mínima TATA BOX) e seqüências adicionais reguladoras. RNA polimerase II liga-se ao promotor mínimo TATA BOX. Esse é o local de união do complexo inicial de transcrição. O promotor mínimo pode ser regulado por seqüências reguladoras e conservadas de DNA. São conhecidas como CAAT BOX e GC BOX São locais de ligações de outras proteínas fatores transcricionais.
 e seqüências adicionais reguladoras. RNA polimerase II liga-se ao promotor mínimo TATA BOX. Esse é o local de união do complexo inicial de transcrição. O promotor mínimo pode ser regulado por seqüências reguladoras e conservadas de DNA. São conhecidas como CAAT BOX e GC BOX. São locais de ligações de outras proteínas fatores transcricionais.")
9
COMPLEXO RNA POLIMERASE + FATORES GERAIS DE TRANSCRIÇÃO

10
Seqüência da ligação dos fatores gerais de transcrição à RNA polimerase II.
Promotor Complexo inicial de transcrição Fosforilação de DNA

11
Fatores transcricionais reguladores

12
CAAT BOX e GC BOX podem reprimir ou ativar a expressão do gene.
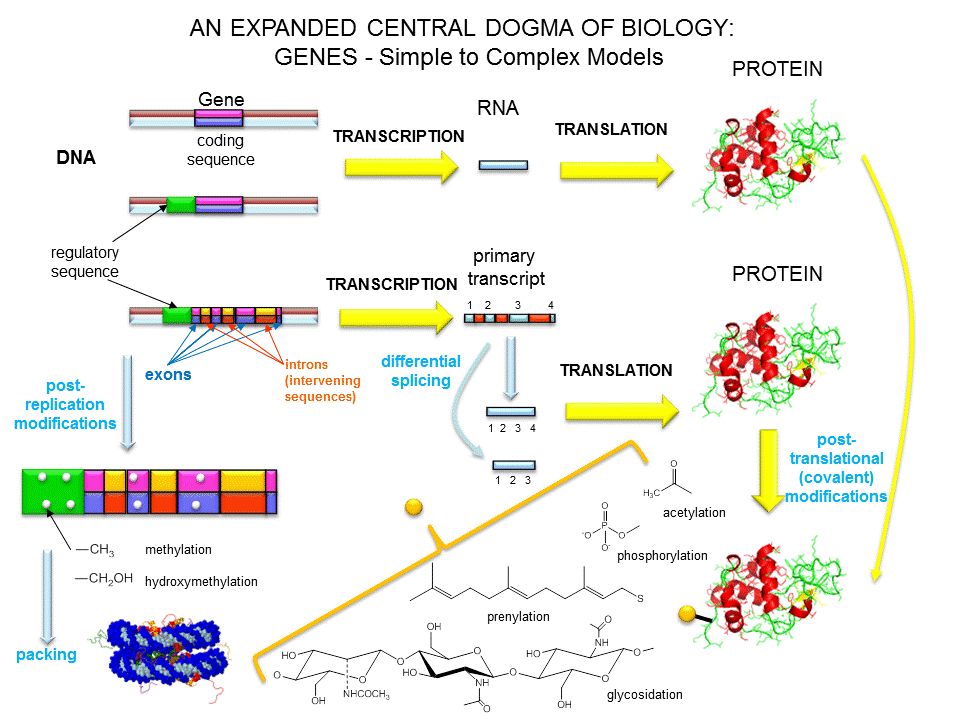
O processo de expressão gênica apresenta duas grandes fases. 1ª FASE - transcrição: síntese de mRNA copiado do molde de DNA. 2ª FASE - tradução: síntese de proteínas nos ribossomos(citoplasma). Genes eucarióticos possuem regiões codificadoras exons e não codificadoras introns. Pré-mRNA sofre processamento com retirada de seqüências introns. A seguir ocorre a tradução no RE.
. Genes eucarióticos possuem regiões codificadoras exons e não codificadoras introns. Pré-mRNA sofre processamento com retirada de seqüências introns. A seguir ocorre a tradução no RE.")
13
Expressão gênica em células eucarióticas
Formação de mRNA com retirado dos introns

14
Expressão gênica em células eucarióticas

15
Regulagem de expressão gênica

16
Regulagem de expressão gênica

17
Regulagem de expressão gênica
Transcrição Tradução

19
Síntese de polipeptídio

20
Códon e anticódon

21
Ribossomo e síntese de polipeptídio

22
Seqüências adicionais de DNA de célula vegetal reguladas por hormônios são chamadas de elementos de resposta. Na transcrição, o DNA forma alças (looping) entre o promotor e seqüências adicionais. Há contato físico entre as seqüências reguladoras havendo controle positivo ou negativo. Após tradução, as proteínas podem durar poucos minutos sendo destruídas. Ubiquitina é um peptídeo pequeno (76aa) que liga-se e marca proteínas a serem destruídas no complexo proteolítico, o proteossomo 26S. Mais de 90% das proteínas têm vida curta.
 entre o promotor e seqüências adicionais. Há contato físico entre as seqüências reguladoras havendo controle positivo ou negativo. Após tradução, as proteínas podem durar poucos minutos sendo destruídas. Ubiquitina é um peptídeo pequeno (76aa) que liga-se e marca proteínas a serem destruídas no complexo proteolítico, o proteossomo 26S. Mais de 90% das proteínas têm vida curta.")
23
Organização e regulação de um gene eucariótico

24
Fundamentos da transdução (tradução + indução) de sinais em plantas
Hormônios vegetais são mensageiros químicos primários. Adaptam crescimento e desenvolvimento da planta às quantidades e qualidades de luz e outros fatores ambientais. Para atuarem, Hormônios (H) ligam-se às proteínas receptoras (R) Forma-se o complexo H + R. Hormônios lipofílicos atravessam membranas celulares e possuem receptores no núcleo, citoplasma, RE. Hormônios hidrossolúveis não entram na célula e possuem receptores na membrana plasmática. Complexo H + R inicia via de transdução de sinais com síntese ou ativação de mensageiros secundários.
 ligam-se às proteínas receptoras (R) Forma-se o complexo H + R. Hormônios lipofílicos atravessam membranas celulares e possuem receptores no núcleo, citoplasma, RE. Hormônios hidrossolúveis não entram na célula e possuem receptores na membrana plasmática. Complexo H + R inicia via de transdução de sinais com síntese ou ativação de mensageiros secundários.")
25
Sinalização transmembrana de esteróides em plantas
Sinalização transmembrana de esteróides em plantas. A molécula do esteróide (em amarelo) se liga ao domínio de ligação do receptor quinase BRI1 (em azul). Isto gera uma plataforma de encaixe para a proteína co-receptora BAK1 (cor de laranja), que é recrutada para o complexo. Interação física entre as BRI1 e BAK1 inicia a resposta de sinalização dentro da célula. Crédito: Zina Deretsky, National Science Foundation
 se liga ao domínio de ligação do receptor quinase BRI1 (em azul). Isto gera uma plataforma de encaixe para a proteína co-receptora BAK1 (cor de laranja), que é recrutada para o complexo. Interação física entre as BRI1 e BAK1 inicia a resposta de sinalização dentro da célula. Crédito: Zina Deretsky, National Science Foundation.")
26
Receptores de hormônios vegetais em geral são proteínas semelhantes as enzimas quinases (ou cinases) e são autofosforiláveis. Quinases são enzimas que tranferem grupos fosfatos de doadores de alta energia, como ATP. Única molécula de hormônio tem seu sinal amplificado. Ocorre ativação enzimática e produção de centenas de moléculas de mensageiros secundários. Os mensageiros secundários mais estudados são: cAMP, DAG, IP3, Ca+2. Hormônios esteroidais entram no núcleo e agem como fatores de transcrição ativando uma proteína receptora. Brassinosteróides são hormônios esteroidais de plantas.

27
Produção de cAMP (adenosina monofosfato cíclica)
Em hormônios hidrossolúveis o complexo H + R forma-se na membrana plasmática. Ativa uma proteína heterotrimérica G presente na membrana. Proteína heterotrimérica G tem três sub-unidades :, , . liga-se a GTP (guanidina trifosfato) que ativa a enzima adenilil-ciclase produzindo cAMP. cAMP ativa enzima quinase A (PKA) que fosforila proteínas fatores de transcrição. Esse modelo ocorre também em plantas. Em plantas, cAMP ativa canais de K+ de membranas celulares de células guardas de Vicia faba. Esse processo ocorre durante a abertura de estômatos.
 que ativa a enzima adenilil-ciclase produzindo cAMP. cAMP ativa enzima quinase A (PKA) que fosforila proteínas fatores de transcrição. Esse modelo ocorre também em plantas. Em plantas, cAMP ativa canais de K+ de membranas celulares de células guardas de Vicia faba. Esse processo ocorre durante a abertura de estômatos.")
28
Modelo de transdução de sinais Síntese de cAMP
H+R ativa proteína G substituindo GDP por GTP Ativação da adenilil-ciclase Síntese de cAMP

29
c AMP

30
Produção de IP3 (inositol-tri-fosfato) e DAG (diacil-glicerol)
Complexo H + R: ativação de proteína G e ativação de fosfolipase C (PLC). PLC hidrolisa o fosfolipídio de membrana PIP2 (fosfatidil –inositol 4,5- bifosfato) Formação de IP3 (inositol-3-fosfato) e DAG (diacil-glicerol). IP3 é hidrossóluvel e difunde-se no citosol. IP3 + receptor do tonoplasto: abre canais de Ca2+ do tonoplasto Isso gera aumento temporário dos níveis de Ca2+ no citosol.
. PLC hidrolisa o fosfolipídio de membrana PIP2 (fosfatidil –inositol 4,5- bifosfato) Formação de IP3 (inositol-3-fosfato) e DAG (diacil-glicerol). IP3 é hidrossóluvel e difunde-se no citosol. IP3 + receptor do tonoplasto: abre canais de Ca2+ do tonoplasto. Isso gera aumento temporário dos níveis de Ca2+ no citosol.")
31
Aumentos de Ca2+ ocorrem em células-guardas de estômatos no mecanismo de fechamento induzido por ABA (ÁCIDO ABSCÍSICO). Ca2+ pode se ligar a uma proteína calmodulina e ativar várias enzimas. Há proteínas quinases dependentes de Ca2+ envolvidas na resposta de fechamento estomático induzida por ABA. Esse processo foi estudado na regulagem de canais de ânions no tonoplasto de células guardas de Vicia faba. DAG (lipídio associado à membrana) ativa proteína quinase C (PKC) que fosforila canais de íons, fatores de transcrição e outras enzimas.
 ativa proteína quinase C (PKC) que fosforila canais de íons, fatores de transcrição e outras enzimas.")
32
Formação de IP3 e DAG

33
Formação de DAG e IP3-Função de IP3

34
CALMODULINA

35
Controle do ciclo celular
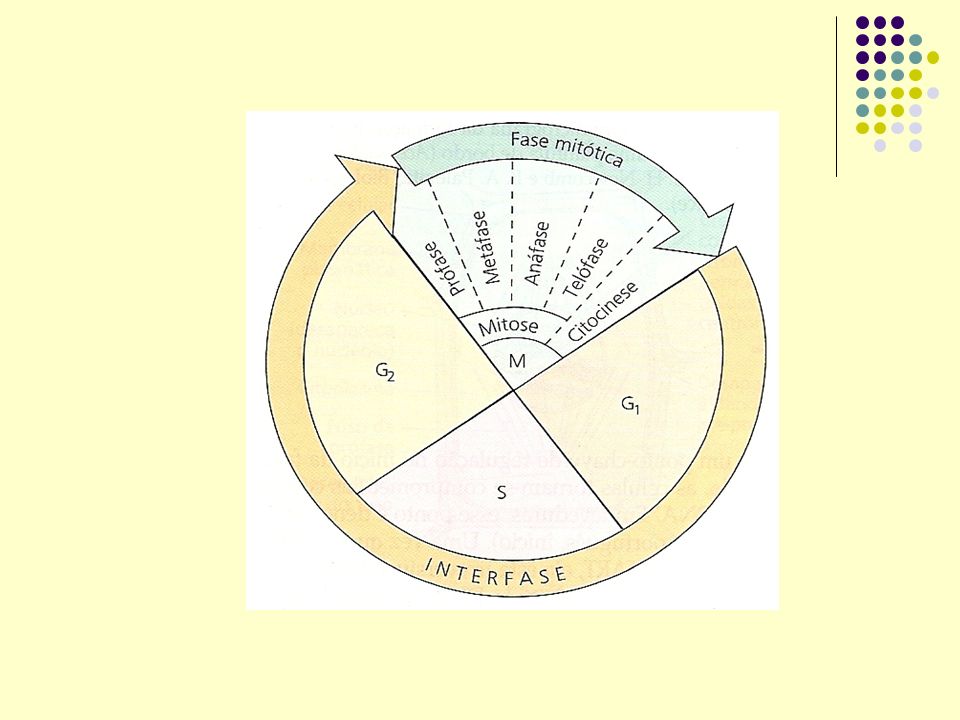
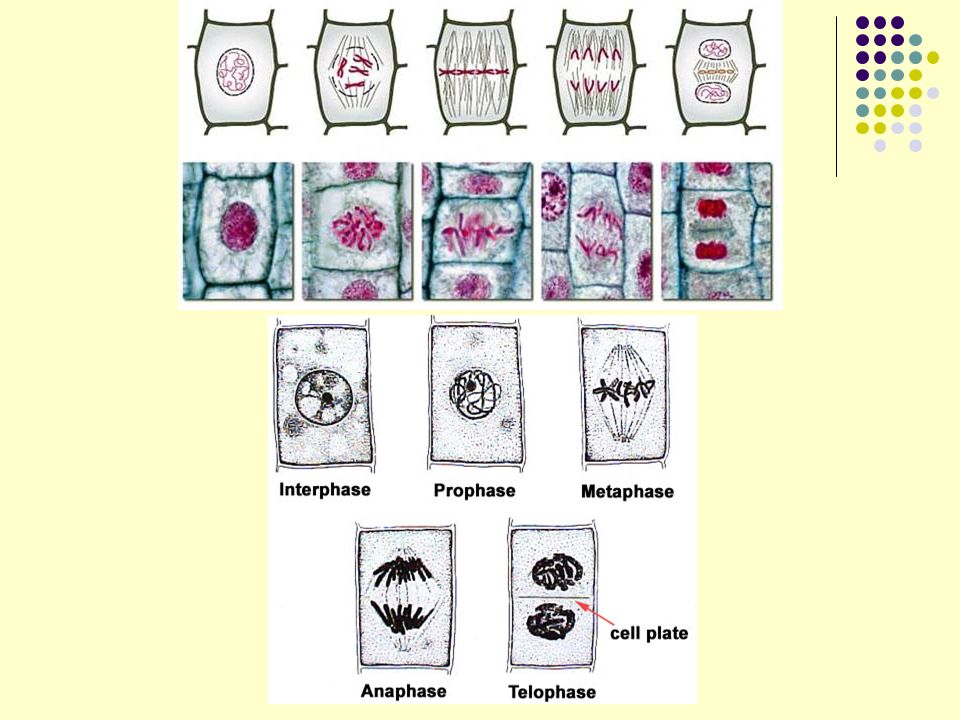
Os hormônios vegetais estão também envolvidos no controle do ciclo celular. Fases do ciclo celular : G1, S e G2 (interfase); M (mitose). G1 é o START: preparação para duplicação de DNA nuclear, pelo complexo de pré-replicação de cromatina. S: DNA replicado. G2 : preparo para mitose. Enzimas que controlam transição do ciclo celular: proteínas quinases dependentes de proteínas ciclinas (CDKS). Hormônios vegetais induzem a expressão gênica de CDKs e ciclinas. CDK fazem transição de G1 para S e de G2 para M (mitose).
; M (mitose). G1 é o START: preparação para duplicação de DNA nuclear, pelo complexo de pré-replicação de cromatina. S: DNA replicado. G2 : preparo para mitose. Enzimas que controlam transição do ciclo celular: proteínas quinases dependentes de proteínas ciclinas (CDKS). Hormônios vegetais induzem a expressão gênica de CDKs e ciclinas. CDK fazem transição de G1 para S e de G2 para M (mitose).")
36
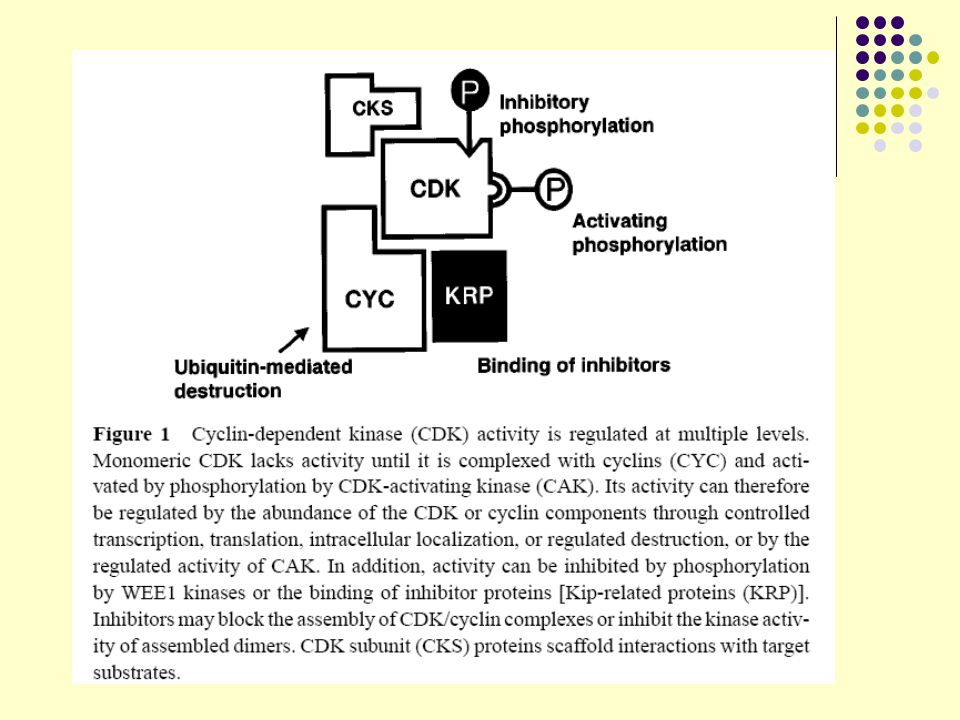
Regulação de atividade de CDKS:
1) Síntese de proteínas ciclinas. 2) Destruição de ciclinas pela ubiquitina no proteossomo 26 S. 3) Fosforilação e desfosforilação de resíduos de aminoácidos na molécula de CDK pelas fosfatases. Mudança de G1 para S necessita de ciclinas TIPO G. Mudança de G2 para M necessita de ciclinas mitóticas. Fosforilação de algumas moléculas de tirosinas das CDKs faz a ativação. Outras moléculas de tirosinas ao se fosforilarem inibem atividade.
 Síntese de proteínas ciclinas. 2) Destruição de ciclinas pela ubiquitina no proteossomo 26 S. 3) Fosforilação e desfosforilação de resíduos de aminoácidos na molécula de CDK pelas fosfatases. Mudança de G1 para S necessita de ciclinas TIPO G. Mudança de G2 para M necessita de ciclinas mitóticas. Fosforilação de algumas moléculas de tirosinas das CDKs faz a ativação. Outras moléculas de tirosinas ao se fosforilarem inibem atividade.")
38
CDK ativa estimula mitose

41
CDKs são inativas até que se complexem com ciclinas.
O complexo é ativado por fosforilação através da enzima CDK quinase (CAK).
.")
43
Referências Dewitte, W &. Murray, J.A.H THE PLANT CELL CYCLE Annu. Rev. Plant Biol :235– Taiz, L. & Zeiger, E Plant Physiology. Sinauer Associates, Inc, Publishers, 792p. Taiz, L. & Zeiger, E Plant Physiology. Sinauer Associates, Inc, Publishers, 705p. Taiz, L. & Zeiger, E Sinauer Associates, Inc, Publishers.

Apresentações semelhantes















 -receptor ê Ativação do receptor do fator de.>")





