Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Territórios Cromossômicos Interfásicos
Genética Humana Molecular Profa. Dra. Ana Elizabete Silva

2
INTRODUÇÃO Organização Hierárquica da Cromatina
Avanços tecnológicos para avaliação dos cromossomos interfásicos Organização do genoma no núcleo interfásico: Territórios Cromossômicos Organização não-casual dos cromossomos interfásicos Dinâmica dos cromossomos Posicionamento preferencial dos genes nos TC Compartimentalização do núcleo interfásico Rearranjos cromossômicos e proximidade espacial

3
Após a era genômica... Como o genoma está funcionalmente organizado e confinado no núcleo? Qual a organização dos cromossomos dentro do núcleo? Há um local preferencial? Como ocorre a dinâmica entre o genoma e seus fatores regulatórios para expressão e silenciamento gênico? Há diferenças no posicionamento de genes ativos x inativos no núcleo?

4
Arranjos dos cromossomos no núcleo interfásico em não-território (superior) e em territórios cromossômicos (inferior)
 e em territórios cromossômicos (inferior)")
5
Genoma Humano Cerca de genes → 3.2 bilhões de pb (2m de comprimento linear na forma despida) → empacotado em estrutura de ordem superior (~10 mil vezes) → volume nuclear μm3 Como os genes estão arranjados no núcleo interfásico (46 cromossomos), para que sejam acessíveis aos reguladores transcricionais (expressão e silenciamento)?
 → empacotado em estrutura de ordem superior (~10 mil vezes) → volume nuclear μm3. Como os genes estão arranjados no núcleo interfásico (46 cromossomos), para que sejam acessíveis aos reguladores transcricionais (expressão e silenciamento)")
6
Níveis de Organização da Cromatina
6 níveis interdependentes de organização da cromatina: o DNA despido ao cromossomo 1º- DNA de dupla hélice enrolado ao redor do core de octâmero de histonas (colar de contas) fibra nucleossômica de 10 nm → empacotamento 7x 2º- enrolamento do nucleossomo para formar a fibra de 30 nm (solenóide) organização mais prevalente no núcleo interfásico → empacotamento 40x Fibra de 30nm forma dobras em domínios de alças (variando de Kb) → base para a regulação da replicação/transcrição As alças são unidas à base de um eixo central (esqueleto do cromossomo) → arranjamento radial das alças 6º- Cromossomos metafásicos: maior nível de compactação da cromatina → empacotamento x
 fibra nucleossômica de 10 nm → empacotamento 7x. 2º- enrolamento do nucleossomo para formar a fibra de 30 nm (solenóide) organização mais prevalente no núcleo interfásico → empacotamento 40x. Fibra de 30nm forma dobras em domínios de alças (variando de Kb) → base para a regulação da replicação/transcrição. As alças são unidas à base de um eixo central (esqueleto do cromossomo) → arranjamento radial das alças. 6º- Cromossomos metafásicos: maior nível de compactação da cromatina → empacotamento x.")
7
Organização Hieráquica da Cromatina
and centromere structure

8
Organização hierárquica da cromatina
H2A H2B H3 H4

9
Era da Citogenética Molecular...
Desenvolvimento de sondas fluorescentes Microscopia fluorescente (análise 2D) Microscopia confocal a laser (análise 3D) Permitiu visualizar aspectos da expressão gênica e dinâmica em alta resolução em células vivas
 Microscopia confocal a laser (análise 3D) Permitiu visualizar aspectos da expressão gênica e dinâmica em alta resolução em células vivas.")
10
Núcleo virtual: A- visualização da cromatina individual B- Imagem de uma secção confocal

11
Organização do genoma no núcleo interfásico
A cromatina descondensada no núcleo interfásico ocupa espaço definido sem sobreposições, separado por domínios intercromossômicos TERRITÓRIOS CROMOSSÔMICOS

12
Territórios Cromossômicos
ESPAÇO SUB-NUCLEAR LIMITADO, EM QUE CROMOSSOMOS INDIVÍDUAIS NO NÚCLEO INTERFÁSICO SÃO OCUPADOS NÃO-CASUALMENTE Proposto por Rabl e Boveri e confirmado por Cremer (1980) Modelo proposto por Cremer et al. (1982): organização de cromossomos individuais em territórios compactos claramente delineados → permite a exposição de sequências expressíveis na superfície territorial. Estes territórios não se sobrepõem, seriam separados por cargas negativas das cromatinas, em pH fisiológico.
 Modelo proposto por Cremer et al. (1982): organização de cromossomos individuais em territórios compactos claramente delineados → permite a exposição de sequências expressíveis na superfície territorial. Estes territórios não se sobrepõem, seriam separados por cargas negativas das cromatinas, em pH fisiológico.")
13
Organização não-casual dos cromossomos interfásicos
HISTÓRICO Carls Rabl (1885): plantas cromossomos ocupam domínios discretos durante intérfase, reflete sua orientação mitótica orientação polarizada da anáfase: permanece até a prófase seguinte Configuração de Rabl: telômeros presos no envelope nuclear de um lado do núcleo e centrômeros no lado oposto (plantas, dípteros, mas não em mamíferos)
: plantas cromossomos ocupam domínios discretos durante intérfase, reflete sua orientação mitótica. orientação polarizada da anáfase: permanece até a prófase seguinte. Configuração de Rabl: telômeros presos no envelope nuclear de um lado do núcleo e centrômeros no lado oposto (plantas, dípteros, mas não em mamíferos)")
14
Orientação polarizada de Rabl no núcleo interfásico:
A- raiz de planta: centrômero (verde); telômeros (vermelho) B- esquema interpretativo: cromossomos na orientação de Rabl com todos centrômeros de um lado do núcleo e todos os telômeros em direção ao lado oposto. Obs.: levedura: cromossomos parecem misturar-se livremente
; telômeros (vermelho) B- esquema interpretativo: cromossomos na orientação de Rabl com todos centrômeros de um lado do núcleo e todos os telômeros em direção ao lado oposto. Obs.: levedura: cromossomos parecem misturar-se livremente. rid=mboc4.figgrp.684.")
15
Organização não-casual dos cromossomos interfásicos
HISTÓRICO década de 40: Alças de ancoragem: heterocromatina constitutiva, facultativa e telômeros unem, na maioria dos casos ao envoltório nuclear.

16
Cromatina Sexual - corpúsculo de Barr

17
Movimentos da cromatina no núcleo interfásico Actina/miosina
Os cromossômos ocupam domínios definidos na interfáse e movem-se muito pouco portanto, a placa metafásica reflete a organização interfásica (orientação polarizada de Rabl durante a intérfase e prófase) Movimentos da cromatina no núcleo interfásico Actina/miosina TFF1: cr. 21 GREB1: cr. 2 Cell. Author manuscript; available in PMC 2010 July 7.
 Movimentos da cromatina no núcleo interfásico. Actina/miosina. TFF1: cr. 21. GREB1: cr. 2. Cell. Author manuscript; available in PMC 2010 July 7.")
18
Conclusões Em geral, os cromossomos, distribuem-se ao acaso (exceção dos Dípteros). Desvios da distribuição metafásica ao acaso podem ser em consequência: - adesividade da heterocromatina; - associação de satélite; e - tendência dos cromossomos pequenos localizar-se no meio da placa metafásica. Em muitos núcleos diferenciados, os centrômeros aderem-se no envoltório nuclear, nos nucléolos ou em ambos

19
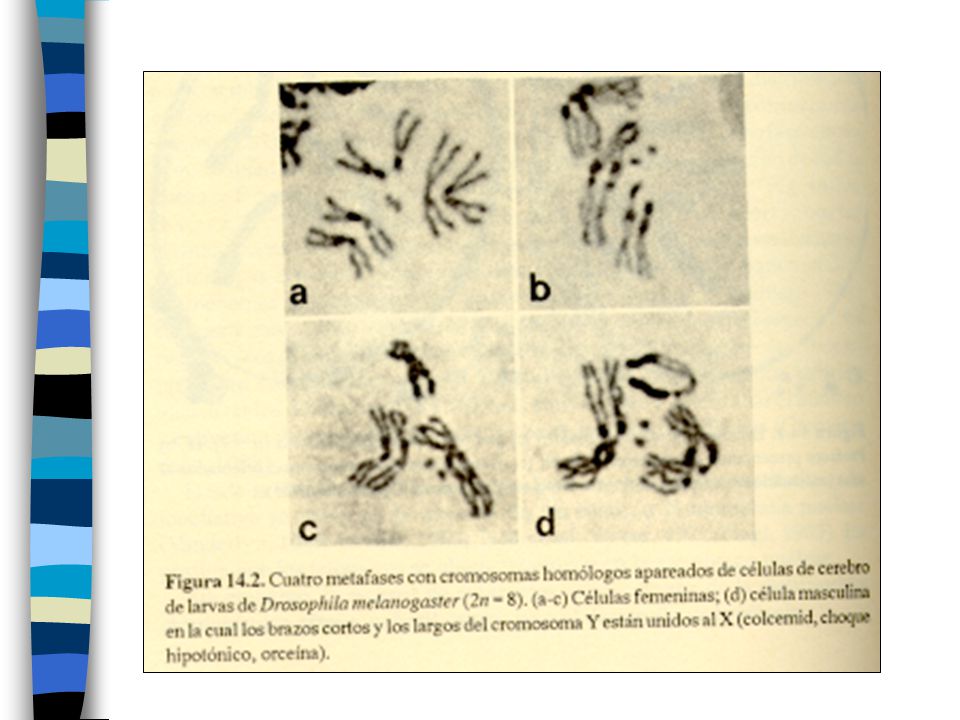
Distribuição Cromossômica Não-Aleatória
Cromossomos homólogos estão pareados durante toda a intérfase? Depende da espécie e do tipo celular Linfócitos e fibroblastos humanos: para os cromossomos grandes é infrequente, enquanto para os cromossomos pequenos denso em genes é mais frequente a associação homóloga de TCs. Mamíferos: os TC homólogos são usualmente não-adjacentes Apenas em Dípteros os homólogos permanecem pareados mesmo em metáfases tratadas com colcemide e solução hipotônica Cada cromossomo une-se em um ponto pré-determinado do envoltório nuclear (poros ou lamina nuclear) originando uma ordem cronológica mais ou menos constante na metáfase.
 originando uma ordem cronológica mais ou menos constante na metáfase.")
21
Posicionamento Radial dos cromossomos em células de mamíferos:
a) Configuração de Rabl: centrômeros (vermelho) e telômeros (amarelos) → alinhados em lados opostos do núcleo b) posicionamento radial dos cromossomos: localização preferencial dos cromossomos na periferia ou interior do núcleo (conforme tamanho ou densidade gênica) c) posicionamento relativo dos TC em posições preferenciais em relação ao outros: cr grandes → periferia do núcleo cr pequenos → interior do núcleo
 Configuração de Rabl: centrômeros (vermelho) e telômeros (amarelos) → alinhados em lados opostos do núcleo. b) posicionamento radial dos cromossomos: localização preferencial dos cromossomos na periferia ou interior do núcleo (conforme tamanho ou densidade gênica) c) posicionamento relativo dos TC em posições preferenciais em relação ao outros: cr grandes → periferia do núcleo. cr pequenos → interior do núcleo.")
22
Território Cromossômico de galinha:
-9 pares de macrocromossomos (pobres em genes-replicação tardia): localizados preferencialmente na periferia do núcleo - 30 pares de microcromossomos (ricos em genes-replicação precoce): interior do núcleo d- cromossomos homólogos em localizações não-adjacentes
: localizados preferencialmente na periferia do núcleo pares de microcromossomos (ricos em genes-replicação precoce): interior do núcleo. d- cromossomos homólogos em localizações não-adjacentes.")
23
TC humanos: (a-c) cr. grandes:1-5,X) (vermelho-periferia); cr
TC humanos: (a-c) cr.grandes:1-5,X) (vermelho-periferia); cr.pequenos:(17-20) (verde-interior) TC galinhas: (d-h) Microcromossomos: replicação precoce (interior do núcleo) Macrocromossomos: replicação tardia (periferia do núcleo) Macrocromossomos homólogos não-adjacentes (periferia) Linfócitos fibroblastos
 cr.grandes:1-5,X) (vermelho-periferia); cr.pequenos:(17-20) (verde-interior) TC galinhas: (d-h) Microcromossomos: replicação precoce (interior do núcleo) Macrocromossomos: replicação tardia (periferia do núcleo) Macrocromossomos homólogos não-adjacentes (periferia) Linfócitos fibroblastos.")
24
Territórios cromossômicos humanos ricos e pobres em genes
TCs com conteúdo de DNA similar mas com densidade gênica muito diferentes: ocupam posição nuclear distinta no exterior ou interior do núcleo → conteúdo de genes é um determinante chave do posicionamento dos TCs. Exemplo: cromossomos 18 e 19 humanos (conteúdo de DNA similar: 85 e 67 Mb) -cr 18 (vermelho) pobre em genes-replicação tardia: periferia -cr 19 (verde) rico em genes -replicação precoce: interior
 -cr 18 (vermelho) pobre em genes-replicação tardia: periferia. -cr 19 (verde) rico em genes -replicação precoce: interior.")
25
LOCALIZAÇÃO DOS TELÔMEROS
-compartilham pequenos territórios dinamicamente associados posicionados preferencialmente na interface dos domínios de cromatina -movimento constrito dentro do TC Cytometry Part A 75A: , 2009

26
Movimento e Associação de Telômeros
Cytometry Part A 75A: , 2009

27
Proposta: cada cromossomo é marcado por um “zip-code” (determinantes) que determina sua posição não casual no núcleo Objetivos: a-investigar se núcleo de camundongo reconhece “determinantes” de posição nuclear de cromossomos humanos b-se o posicionamento cromossômico e atividade transcricional permanecem acopladas nessas condições trans-espécies

28
Núcleos de linhagem celular A9- fibroblastos híbridos camundongo-humano
vermelho: cr. 7 (pobre em genes) - periferia nuclear Amarelo: cr. 18 (pobre em genes) – periferia nuclear Verde: cr. 19 (rico em genes) – centro nuclear -o posicionamento dos cromossomos humanos pobres e ricos em genes é conservado em núcleos de camundongo. -estudo de perfil gênico dos 3 crs. humanos demonstrou que são transcricionalmente ativos nas células de camundongo
 - periferia nuclear. Amarelo: cr. 18 (pobre em genes) – periferia nuclear. Verde: cr. 19 (rico em genes) – centro nuclear. -o posicionamento dos cromossomos humanos pobres e ricos em genes é conservado em núcleos de camundongo. -estudo de perfil gênico dos 3 crs. humanos demonstrou que são transcricionalmente ativos nas células de camundongo.")
29
Verde: 19p Vermelho: 19q del(19q) : análise de expressão gênica evidenciou apenas expressão de genes de 19p Conclusões: cromossomos humanos introduzidos artificialmente em células são transcricionalmente ativos no genoma de camundongo. Portanto a densidade gênica/atividade transcricional de um cromossomo serve como “determinante” para a posição do TC nos núcleos de mamíferos.

30
O CENTRO NUCLEAR É MAIS PROPÍCIO PARA A TRANSCRIÇÃO
CONCLUSÕES O posicionamento radial dos cromossomos depende: Tamanho do cromossomo: conteúdo de DNA (pb) Densidade gênica/atividade transcricional Tempo de replicação (precoce x tardia) Etapas do ciclo celular e tipos celulares diferentes O CENTRO NUCLEAR É MAIS PROPÍCIO PARA A TRANSCRIÇÃO
 Densidade gênica/atividade transcricional. Tempo de replicação (precoce x tardia) Etapas do ciclo celular e tipos celulares diferentes. O CENTRO NUCLEAR É MAIS PROPÍCIO PARA A TRANSCRIÇÃO.")
31
Contudo, há exceções... Há estudos mostrando genes ativos na periferia nuclear: -Saccharomyces cerevisae: genes associados com complexo do poro nuclear -expressão do gene -globina iniciada na periferia movimento p/interior do núcleo durante maturação eritróide

32
Dinâmica dos Cromossomos
Células fixadas: TCs ocupam região nuclear discreta, mas TCs são dinâmicos em núcleos interfásicos: posição pode ser dependente do ciclo celular Células G1: centrômero do cr 8: localizado adjacente a periferia nuclear e braços cromossômicos estendidos em direção ao interior do núcleo Células G2: reorientação do cromossomo: centrômero posicionado interiormente e braços estendidos em direção a periferia nuclear

33
Dinâmica dos Cromossomos
Células vivas: análise do ciclo celular: movimento de sítios marcados em direção ao interior nuclear relacionado com replicação do DNA região nuclear mais interna, mais favorável para locos potencialmente ativos Células vivas: TCs mostram apenas pequenos movimentos; células neuronais: observado grandes movimentos da cromatina durante a diferenciação, assim como em Drosophila durante a interfase Leveduras: núcleos em G1: movimentos grandes rápidos dependentes de ATP, nos pontos de origem de replicação ação de enzimas dependentes de ATP, envolvidas na transcrição e remodelamento da cromatina

34
Early S replicating chromatin domains
Mid (Late) S replicating chromatin domains 3-5 cell divisions 3-5 divisões celulares Domínio de cromatina de replicação precoce Domínio de cromatina de replicação –média-tardia Medida comparativa do movimento dos domínios da cromatina dentro dos TC individuais em células vivas: Após algumas gerações com marcação é possivel distinguir TC individuais TCs não são estáticos cromatina e componentes não cromatínicos no núcleo interfásico podem mudar em associação com alterações na função celular potencialmente dirigidas por proteínas nucleares motoras como acto-miosina
 S replicating chromatin domains. 3-5 cell divisions. 3-5 divisões celulares. Domínio de cromatina de replicação precoce. Domínio de cromatina de replicação –média-tardia. Medida comparativa do movimento dos domínios da cromatina dentro dos TC individuais em células vivas: Após algumas gerações com marcação é possivel distinguir TC individuais. TCs não são estáticos cromatina e componentes não cromatínicos no núcleo interfásico podem mudar em associação com alterações na função celular potencialmente dirigidas por proteínas nucleares motoras como acto-miosina.")
35
Manutenção da Posição dos Cromossomos Interfásicos
TC células vivas: movimento de translação limitado (G1 tardio, S e G2): a-Restrição do volume nuclear; Retenção física: b-interação da cromatina c/componentes da lâmina ou do envelope nuclear c-ligação c/estruturas nucleares internas: lâmina interna (esqueleto nuclear); associação c/nucléolo e compartimentos de fatores splicing do pré-mRNA. INM(membrana nuclear interna)-compartimento da lamina nuclear: a- em células de mamíferos: atua no estabelecimento e manutenção da cromatina transcricionalmente inativa, b- leveduras: a periferia nuclear atua tanto no silenciamento (repressão dos telômeros) como ativação gênica (associação com complexos do poro nuclear-NPC)
: a-Restrição do volume nuclear; Retenção física: b-interação da cromatina c/componentes da lâmina ou do envelope nuclear. c-ligação c/estruturas nucleares internas: lâmina interna (esqueleto nuclear); associação c/nucléolo e compartimentos de fatores splicing do pré-mRNA. INM(membrana nuclear interna)-compartimento da lamina nuclear: a- em células de mamíferos: atua no estabelecimento e manutenção da cromatina transcricionalmente inativa, b- leveduras: a periferia nuclear atua tanto no silenciamento (repressão dos telômeros) como ativação gênica (associação com complexos do poro nuclear-NPC)")
36
Objetivos: estudar as consequências da atividade gênica após compartimentalização direcionada p/INM-lamina, usando um segmento cromossômico contendo sítio de ligação à proteínas da lamina nuclear e um gene reporter. -Gene pode ser reposicionado p/ INM-lamina e sofrer repressão como consequência dessa compartimentalização? Ou -Gene pode sofrer repressão no interior do núcleo e então ser reposicionado p/INM-lamina para estabilizar a estrutura da heterocromatina?

37
Sistema para compartimentalização direcionada de região genômica:
-Resultados demonstraram que recrutamento de genes alvos p/INM-lamina resulta em repressão transcricional Modelo proposto: INM-lamina reprime a atividade gênica até que um transativador potente seja capaz de sobrepor a repressão imposta pela interação com este compartimento. -Vários genes associados a INM-lamina são transcricionalmente inativos

38
-estado não amarrado: lacO posicionado centralmente no núcleo
-estado amarrado: lacO posicionado na borda do núcleo Conclusão: compartimentalização nuclear parece ter uma função importante na regulação gênica INM-lamina reprime a atividade gênica Ex.: vários genes do desenvolvimento associados a INM-lamina quando transcricionalmente inativos

39
Posicionamento preferencial dos genes nos TC
Há alguma relação entre posição do gene no TC com atividade gênica? Alguns estudos têm sugerido: Genes inativos: localizados no interior das regiões do TC Genes ativos: concentram na periferia do TC → adjacente ao espaço nucleoplásmico → Domínio do Compartimento Intercromossômico (ICD)
")
40
Modelo ICD: pré-mRNA nascente localiza-se na superfície dos domínios cromossômicos
Os TCs são separados por canais intercromatínicos (retículo contínuo de espaço livre de DNA e possivelmente ocupado por matriz nuclear e outros fatores não-cromatínicos funcionalmente ativos) tais canais seriam contínuos com os poros nucleares e representariam sítios de processamento pós-transcricional direcional, culminando na passagem núcleo-citoplasma.
 tais canais seriam contínuos com os poros nucleares e representariam sítios de processamento pós-transcricional direcional, culminando na passagem núcleo-citoplasma.")
41
ICD IC CT Modelo de Domínio Intercromossomo (ICD) x Modelo de Compartimento Intercromatina (IC) Modelo ICD: genes ativos (verde) → localizados na superfície dos TC → acessíveis aos fatores de transcrição/splicing do ICD → transcritos facilmente transportados para os poros nucleares. Genes inativos ou locos intergênicos (vermelho) → encontrados no interior dos TC → inacessíveis aos fatores do ICD. Modelo IC: espaço de não-cromatina e fatores → localizam dentro dos TC Genes ativos: superfície dos territórios ou superfície da cromatina condensada; Genes inativos: dentro da cromatina condensada
 → localizados na superfície dos TC → acessíveis aos fatores de transcrição/splicing do ICD → transcritos facilmente transportados para os poros nucleares. Genes inativos ou locos intergênicos (vermelho) → encontrados no interior dos TC → inacessíveis aos fatores do ICD. Modelo IC: espaço de não-cromatina e fatores → localizam dentro dos TC. Genes ativos: superfície dos territórios ou superfície da cromatina condensada; Genes inativos: dentro da cromatina condensada.")
42
Genes ativos: encontrados no interior e periferia do TC
Regiões transcricionalmente ativas: encontradas em alças extendidas para fora do TC Regiões não codificadoras e genes inativos (vermelho): podem ser encontradas na periferia ou interior do TC Cr 12 (vermelho); 14 (amarelo); 15 (verde): linfócitos de camundongo b-c) Intérfase: TC em fibroblastos de camundongo d) Genes ativos x inativos
: podem ser encontradas na periferia ou interior do TC. Cr 12 (vermelho); 14 (amarelo); 15 (verde): linfócitos de camundongo. b-c) Intérfase: TC em fibroblastos de camundongo. d) Genes ativos x inativos.")
43
Associação com densidade gênica e expressão gênica.
Exemplos: EDC: 1q21 (>40genes)→ localizado externamente no TC 1 ou em grandes alças: durante diferenciação de queratinócitos MHC: 6p21.3 → localizado externamente no TC 6 ou em grandes alças: principalmente em células B-linfoblastóides e fibroblastos após tratamento com interferon Associação com densidade gênica e expressão gênica.
→ localizado externamente no TC 1 ou em grandes alças: durante diferenciação de queratinócitos. MHC: 6p21.3 → localizado externamente no TC 6 ou em grandes alças: principalmente em células B-linfoblastóides e fibroblastos após tratamento com interferon. Associação com densidade gênica e expressão gênica.")
44
Modelo funcional da arquiterura nuclear:
a)TC: alça cromatínica genes ativos; b)TC: domínios separados para os braços curto e longo e centrômero (*). Genes (branco) ativamente transcritos → localizados na alça cromatínica. Recrutamento deles (preto) para a heterocromatina centromérica → silenciamento; c)TC densidade cromatínica variável (laranja: alta densidade; amarelo: baixa densidade); d)TC: domínio de replicação precoce (verde) domínio de replicação média e tardia (vermelho). Cromatina pobre em genes→periferia nuclear próxima a lâmina nuclear (amarela), nas dobras da lâmina e redor do nucléolo.Cromatina rica em genes → localizada entre os compartimentos pobre em genes.e)estrutura da cromatina de ordem superior: genes ativos (branco)→superfície da fibra cromatínica, genes silenciados (preto) interior; f) modelo IC: o IC (verde) contém complexos (pontos laranjas) e grandes domínios não cromatínicos (agregações dos pontos laranjas) para transcrição, splicing, replicação e reparo; g) TC com domínio cromatínico de ~1=Mb (vermelho) e IC (verde) expandindo entre esses domínios
TC: alça cromatínica genes ativos; b)TC: domínios separados para os braços curto e longo e centrômero (*). Genes (branco) ativamente transcritos → localizados na alça cromatínica. Recrutamento deles (preto) para a heterocromatina centromérica → silenciamento; c)TC densidade cromatínica variável (laranja: alta densidade; amarelo: baixa densidade); d)TC: domínio de replicação precoce (verde) domínio de replicação média e tardia (vermelho). Cromatina pobre em genes→periferia nuclear próxima a lâmina nuclear (amarela), nas dobras da lâmina e redor do nucléolo.Cromatina rica em genes → localizada entre os compartimentos pobre em genes.e)estrutura da cromatina de ordem superior: genes ativos (branco)→superfície da fibra cromatínica, genes silenciados (preto) interior; f) modelo IC: o IC (verde) contém complexos (pontos laranjas) e grandes domínios não cromatínicos (agregações dos pontos laranjas) para transcrição, splicing, replicação e reparo; g) TC com domínio cromatínico de ~1=Mb (vermelho) e IC (verde) expandindo entre esses domínios.")
45
Diferentes modelos da arquitetura nuclear:
A- Modelo CT-IC (Chromosome Territory-Interchromatin Compartment) B- Arquitetura nuclear conforme modelo CT-IC (PF:fibrila pericromatina): genes parcialmente descondensados RNA nascente (verde) e RNA pol (branco) C- pontos em azul (contatos intra e intercromossomal) D- colocalização dos genes para expressão (pontos rosas: fatores de transcrição podem recrutar genes; circulo azul: corregulação in trans; área rosa: speckle) Cite this article as Cold Spring Harb Perspect Biol 2010;2:a003889
 B- Arquitetura nuclear conforme modelo CT-IC (PF:fibrila pericromatina): genes parcialmente descondensados RNA nascente (verde) e RNA pol (branco) C- pontos em azul (contatos intra e intercromossomal) D- colocalização dos genes para expressão (pontos rosas: fatores de transcrição podem recrutar genes; circulo azul: corregulação in trans; área rosa: speckle) Cite this article as Cold Spring Harb Perspect Biol 2010;2:a")
46
Objetivos: análise da posição dos genes CCND1 (TC 11) e HER-2/neu (TC 17) nos respectivos TCs em câncer de esôfago e gástrico e relação com amplificação gênica e expressão das proteínas
 e HER-2/neu (TC 17) nos respectivos TCs em câncer de esôfago e gástrico e relação com amplificação gênica e expressão das proteínas")
47
CCND1 ou HER-2/neu (vermelho)
Frequências dos TCs com genes localizados na periferia/externo: Tumores esôfago: 61% e 65% dos TCs Mucosa normal: 56% e 33% dos TCs Tumores gástricos: 84% e 67% dos TCs Mucosa normal: 50% e 59% dos TCs Não foi observada associação entre a localização dos genes nos TCs com amplificação e expressão das proteínas

48
Territórios cromossômicos em humanos:
Genes ANT2 e ANT3: exterior de ambos TC Xa ANT2(Xq24): submetido à inativação do X → ativo no Xa e inativo no Xi ANT3(PAR): escapa da inativação no Xi (periferia de ambos Xa e Xi) Territórios cromossômicos em humanos: a-b) braços cromossômicos ocupam CT distintos c-d) bandas com genes ativos ocupam a periferia dos territórios dos cromossomos Xa e Xi e) ANT2 inativo no Xi encontra-se no interior do TC ANT2 ativo no Xa na periferia do TC
: submetido à inativação do X → ativo no Xa e inativo no Xi. ANT3(PAR): escapa da inativação no Xi (periferia de ambos Xa e Xi) Territórios cromossômicos em humanos: a-b) braços cromossômicos ocupam CT distintos. c-d) bandas com genes ativos ocupam a periferia dos territórios dos cromossomos Xa e Xi. e) ANT2 inativo no Xi encontra-se no interior do TC. ANT2 ativo no Xa na periferia do TC.")
49
Dinâmica da Relação Estrutura-Função no Núcleo Interfásico
NUCLÉOLO CROMATINA COMPONENTES NÃO-CROMATÍNICOS COMPARTIMENTALIZADO EM REGIÕES SUB-NUCLEARES DISTINTAS COM FATORES MACROMOLECULARES ENVOLVIDOS EM PROCESSOS NUCLEARES ESPECÍFICOS COMPARTIMENTALIZAÇÃO DINÂMICA

50
NÚCLEO E COMPARTIMENTOS NUCLEARES
Repressão Gênica: -Membrana nuclear interna -Lamina nuclear -Cromatina peri-nucleolar Expressão Gênica: -Complexo do poro nuclear -Corpúsculo PML -Speckles Outros: -Corpúsculo de Cajal -Nucléolo Curr Opin Genet Dev. Author manuscript; available in PMC 2010 April 1.

51
Compartimento Intercromatínico - IC
Espaço entre os territórios cromossômicos: com rede de ribonucleoproteínas e alças cromatínicas - corpúsculos PML (funcionais na leucemia mielóide promielocítica): sítio nuclear dos quais o genoma viral iniciam o centro de replicação viral →processos celulares: defesa viral, supressão tumoral, reparo DNA e regulação transcricional -speckles:regiões formato irregular formada por grânulos intercromatina, ricas em fatores de splicing pré-mRNA - corpúsculos de Cajal:organelas nucleares de função desconhecida → biogênese de snRNP Os TCs têm estrutura complexa dobrada em contato com os canais IC → penetram no interior do TC e permitem a transcrição de genes no interior do TC
: sítio nuclear dos quais o genoma viral iniciam o centro de replicação viral →processos celulares: defesa viral, supressão tumoral, reparo DNA e regulação transcricional. -speckles:regiões formato irregular formada por grânulos intercromatina, ricas em fatores de splicing pré-mRNA. - corpúsculos de Cajal:organelas nucleares de função desconhecida → biogênese de snRNP. Os TCs têm estrutura complexa dobrada em contato com os canais IC → penetram no interior do TC e permitem a transcrição de genes no interior do TC.")
52
DOMÍNIOS NUCLEARES E EXPRESSÃO GÊNICA
...expressão gênica não é influenciada apenas pelo seu compartimento nuclear, mas também por processos dinâmicos: sinalização celular, interação cromossômica, microambiente... Curr Opin Genet Dev. Author manuscript; available in PMC 2010 April 1.

53
Regiões ricas em GC (vermelho); pobres em GC (azul)
M.O. Scheuermann et al. / Experimental Cell Research 301 (2004) 266–279 Regiões ricas em GC (vermelho); pobres em GC (azul)
 266–279. Regiões ricas em GC (vermelho); pobres em GC (azul)")
54
Rearranjos Cromossômicos e Proximidade Espacial
A proximidade espacial de locos gênicos no núcleo contribui para uma maior ocorrência de translocações balanceadas? Por que são mais frequentes alguns tipos de translocações em alguns tipos celulares, como no câncer? Exemplos: linfomas de células B, leucêmias Linfoma de Burkitt: t(8;14)(q24;q21), c-MYC justaposto próximo à cadeia pesada da IGH; IGL(22q11) ou IGK(2p11) Leucêmia meilóide crônica: t(9;22), ABL-BCR
(q24;q21), c-MYC justaposto próximo à cadeia pesada da IGH; IGL(22q11) ou IGK(2p11) Leucêmia meilóide crônica: t(9;22), ABL-BCR.")
55
Efeito da proximidade dos cromossomos nas frequências de translocações:
-determinada, em parte: - organização espacial, em que alguns locos gênicos estão preferencialmente espacialmente próximos em tipos celulares específicos

56
A-Proximidade espacial de translocação preferencial: linhagem dic(3) e t(8;17) resultando em t(dic3;8;17) B-posição relativa do dic(3) e t(8;17): associados significantemente (67% das células); cr. 3 e 8 (72%) e cr. 3 e 19 (40%), apesar da linhagem apresentar 4 cópias do cr. 19.
 e t(8;17): associados significantemente (67% das células); cr. 3 e 8 (72%) e cr. 3 e 19 (40%), apesar da linhagem apresentar 4 cópias do cr. 19.")
57
Posicionamento radial não-casual dos locos em células B normais:
interior mediana periferia Posicionamento radial não-casual dos locos em células B normais: MYC, BCL e IGH: preferencialmente posicionados em proximidade espacial -IGL e CCND1 (interior nuclear) IGH (mediana) BCL6, MYC e IGK (periferia nuclear)
 IGH (mediana) BCL6, MYC e IGK (periferia nuclear)")
58
Proximidade espacial de locos propensos a translocação:
-MYC:IGH → frequentemente próximos em núcleos de linfoma de Burkitt -MYC:TGFBR2 → controle negativo (distantes)
")
59
Sumário Configuração de Rabl não é encontrada em núcleos de células humanas A cromatina não está organizada casualmente dentro do núcleo ocupa TERRITÓRIOS distintos Os territórios dos cromossomos homólogos são usualmente não adjacentes Posição radial dos cromossomos conforme densidade gênica: ricos em genes →interior do núcleo; pobres em genes → direção à periferia Localização estabelecida precocemente no ciclo celular e mantida durante a intérfase

60
Sumário Movimento restrito dos TC durante a interfáse
Genes: movimentos mais dinâmicos na replicação ou transcrição Reposicionamento de genes inativos p/centrômeros → silenciamento Posicionamento de genes ativos no TC não restrita à periferia, devido os domínios intercromatina (IC) IC: abrigam fatores splicing (speckles), maquinaria de transcrição e corpúsculos nucleares (PML e Cajal) Posicionamento relativo não-casual de TC específicos no núcleo: relação com certas translocações mais frequentes
 IC: abrigam fatores splicing (speckles), maquinaria de transcrição e corpúsculos nucleares (PML e Cajal) Posicionamento relativo não-casual de TC específicos no núcleo: relação com certas translocações mais frequentes.")
Apresentações semelhantes