Carregar apresentação
A apresentação está carregando. Por favor, espere

2
FIGURA 3.23Estrutura primária da pró-insulina humana, insulina e peptídeo C. Na pró-insulina, o peptídeo da cadeia-B estende-se da Phe da posição 1 até a Thr da posição 30, o peptídeo C da Arg da posição 31 até a Arg da posição 65, e o peptídeo-A da Gly da posição 66 até a Asn da posição 86. Ligações cistina das posições 7 para 72, 19 para 85 e 71 para 76 são encontradas na pró-insulina.Redesenhado de Bell, G. I., Swain, W. F., Pictet, R., Cordell, B., Goodman, H. M., e Rutter, W. J. Nature 282: 525, 1979.

3
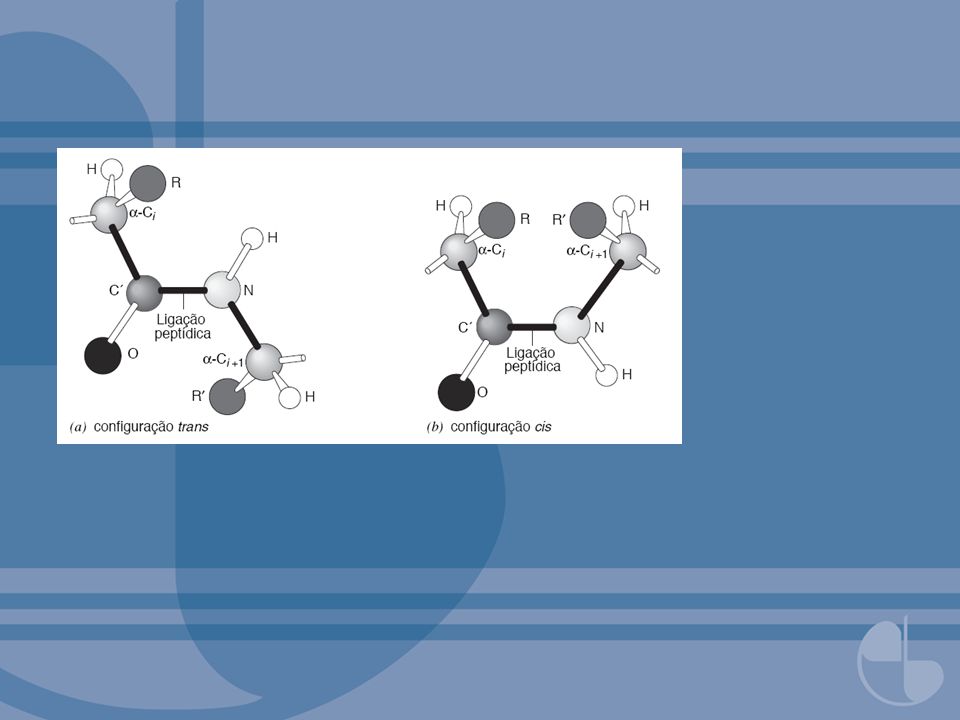
FIGURA 3.25 Passo da hélice (p) para uma hélice com n = 4. Cada círculo sobre uma linha representa um carbono-α de um resíduo de aminoácido. O avanço por resíduo seria p/n De Dickerson, R. E. e Geis, I. The Structural and Action of Proteins. Menlo Park, CA: Benjamin, 1969, p.26. FIGURA 3.24 Cadeia polipeptídica mostrando ligações φ, ψ e peptídicas para o resíduo Ri numa cadeia polipeptídica. Redesenhada com permissão de Dickerson, R. E. e Geis, I. The Structural and Action of Proteins. Menlo Park, CA: Benjamin, 1969, p.25.

4
FIGURA 3.26 Uma α-hélice. Redesenhada com permissão, baseado na figura de Pauling, L. The Nature of Chemical Bond, 3rd ed. Ithanca, NY: Cornell University Press, 1960.

5
FIGURA 3. 27Duas cadeias polipeptídicas em conformação folha-β
FIGURA 3.27Duas cadeias polipeptídicas em conformação folha-β. Cadeias polipeptídicas podem ser adicionadas para gerar estruturas mais extensas.Redesenhada com permissão de Fersht, A. Enzyme Structure and Mechanism, San Francisco: Freeman, 1977, p.10.

6
FIGURA 3.28Exemplo de folha-β antiparalela [resíduos (superior), (meio) e (inferior) da Cu, Zn superóxido dismutase). Linhas tracejadas mostram pontes de hidrogênio entre átomos de oxigênio carbonil (pretos) e átomos de nitrogênio da ligação peptídica (brancos); setas mos-tram direção do polipeptídeo do N-terminal para o C-terminal. Na folha-β característica antiparalela, pares de pontes de hidrogênio intercadeias muito próximas se alternam com pares de pontes de hidrogênio bem espaçadas.Redesenhada com permissão de Richardson, J. S. Adv. Protein Chem. 34: 168, 1981.
 e átomos de nitrogênio da ligação peptídica (brancos); setas mos-tram direção do polipeptídeo do N-terminal para o C-terminal. Na folha-β característica antiparalela, pares de pontes de hidrogênio intercadeias muito próximas se alternam com pares de pontes de hidrogênio bem espaçadas.Redesenhada com permissão de Richardson, J. S. Adv. Protein Chem. 34: 168,")
7
FIGURA 3. 30Estrutura terciária da tripsina
FIGURA 3.30Estrutura terciária da tripsina. (a) Estrutura em fita delineia conformação da cadeia polipeptídica. (b) Estrutura mostra cadeias laterais incluindo resíduos do sítio ativo (marcado com um círculo) com delineamento da cadeia polipeptídica sobreposta (fita). (c) Estrutura de preenchimento de espaço na qual cada átomo é representado com o tamanho de seu raio de van der Waals. Átomos de hidrogênio não são mostrados. Diferentes domínios são mostrados em tons mais escuros e mais claros. Resíduos do sítio ativo estão em cinza com tracejado, e as pontes dissulfeto de cistina intracadeia, em preto circulado de branco. Esferas cinza-claras representam moléculas de água associadas com a proteína. Essa estrutura mostra a densidade de empacotamento no interior da proteína
 Estrutura em fita delineia conformação da cadeia polipeptídica. (b) Estrutura mostra cadeias laterais incluindo resíduos do sítio ativo (marcado com um círculo) com delineamento da cadeia polipeptídica sobreposta (fita). (c) Estrutura de preenchimento de espaço na qual cada átomo é representado com o tamanho de seu raio de van der Waals. Átomos de hidrogênio não são mostrados. Diferentes domínios são mostrados em tons mais escuros e mais claros. Resíduos do sítio ativo estão em cinza com tracejado, e as pontes dissulfeto de cistina intracadeia, em preto circulado de branco. Esferas cinza-claras representam moléculas de água associadas com a proteína. Essa estrutura mostra a densidade de empacotamento no interior da proteína.")
8
FIGURA 3. 31Domínios globulares em proteínas
FIGURA 3.31Domínios globulares em proteínas. (a) Fosfoglicerato quinase tem dois domínios com um braço re-lativamente estreito entre eles. (b) Elastase tem dois domínios fortemente associados separados por uma fenda estreita. Cada esfera no desenho de espaço preenchido representa a posição do carbono-α para um aminoácido na estrutura da proteína.Reimpresso com permissão de Richardson, J. S. Adv. Protein Chem. 34: 168, 1981.
 Fosfoglicerato quinase tem dois domínios com um braço re-lativamente estreito entre eles. (b) Elastase tem dois domínios fortemente associados separados por uma fenda estreita. Cada esfera no desenho de espaço preenchido representa a posição do carbono-α para um aminoácido na estrutura da proteína.Reimpresso com permissão de Richardson, J. S. Adv. Protein Chem. 34: 168,")
9
FIGURA 3.32Desenhos de (a) forma não-ligada da hexoquinase e glicose livre e (b) conformação da hexoquinase com glicose ligada. Nesse desenho de espaço preenchido, cada círculo representa raio de van der Waals de um átomo da estrutura. Glicose é preta, e cada domínio é diferentemente sombreado.Reimpresso com permissão de Bennet, W. S. e Huber, R.. CRC Rev. Biochem. 15:291, 1984.

11
FIGURA 3.35Exemplo de uma dobra domínio-α/β no qual fitas-β formam uma folha-β torcida clássica em lactato desidrogenase e fosfoglicerato quinase. Ver legenda da Figura Como na dobra domínio-α/β anterior, regiões de α-hélice alternam-se com regiões de fita-β na cadeia polipeptídica. Estrutura em folha-β fica no lado de dentro, enquanto os segmentos de α-hélice ficam no lado de fora. Fitas-� ficam em paralelo na estrutura-β.Redesenhado com permissão de Richardson, J. S. Adv. Protein Chem. 34: 168, 1981. FIGURA 3.36Exemplos de superfolds domínio-todo-β: o barril chave grega e dobras jelly roll apresentadas em superóxido dismutase e concanavalina A (ver legenda da Figura 3.33). Fitas-β são principalmente antiparalelas em dobras domínio-todo-β.Redesenhado com permissão de Richardson, J. S. Adv. Protein Chem. 34: 168, 1981.
. Fitas-β são principalmente antiparalelas em dobras domínio-todo-β.Redesenhado com permissão de Richardson, J. S. Adv. Protein Chem. 34: 168,")
12
FIGURA 3.38Diagrama do colágeno demonstrando necessidade de glicinas a cada terceiro resíduo para permitir que diferentes cadeias fiquem muito próximas na estrutura. (a) Modelo em fita para a estrutura superenrolada do colágeno com cada cadeia individual em hélice poliprolina tipo II. (b) Modelo mais detalhado da conformação superenrolada. Todos os átomos de carbono-α são numerados e pontes de hidrogênio propostas são mostradas por linhas tracejadas.Redesenhado com permissão de Dickerson, R. E. e Geis, I. The Structural and Action of Proteins. Menlo Park, CA: Benjamin, 1969, pp. 41, 42.
 Modelo em fita para a estrutura superenrolada do colágeno com cada cadeia individual em hélice poliprolina tipo II. (b) Modelo mais detalhado da conformação superenrolada. Todos os átomos de carbono-α são numerados e pontes de hidrogênio propostas são mostradas por linhas tracejadas.Redesenhado com permissão de Dickerson, R. E. e Geis, I. The Structural and Action of Proteins. Menlo Park, CA: Benjamin, 1969, pp. 41, 42..")
13
FIGURA 3.41Interação de bordas apolares de duas cadeias em conformação em α-hélice em queratina e tropomiosina. Interação de resíduos apolares d-a e a’-d de duas α-hélices alinhadas paralelas na direção NH2-terminal (superior) para COOH-terminal é apresentada.Redesenhado de McLachlan, A. D., e Stewart, M. J. Mol. Biol. 98: 293, 1975 FIGURA 3.43Estrutura geral de lipoproteínas plasmáticas. (a) Modelo da partícula esférica consistindo de núcleo de triacilagliceróis (“E”s cinza bem claro) e ésteres de colesterol (gotinhas mais escuras) com uma capa de ~20 Å de apoliproteínas (indicadas por letras), fosfolipídeos e colesterol não-esterificado. Apoliproteínas ficam embebidas com suas bordas hidrofóbicas orientadas em direção ao núcleo e suas bordas hidrofílicas, em direção ao lado externo. (b) Partícula LDL mostrando ApoB-100 mergulhada na capa externa da partícula.Parte (a) de Segrest, J. P., et al. Adv. Protein. Chem. 45: 303, Parte (b) de Schumaker, V. N., et al., Protein Chem. 45: 205, 1994.
 Modelo da partícula esférica consistindo de núcleo de triacilagliceróis ( E s cinza bem claro) e ésteres de colesterol (gotinhas mais escuras) com uma capa de ~20 Å de apoliproteínas (indicadas por letras), fosfolipídeos e colesterol não-esterificado. Apoliproteínas ficam embebidas com suas bordas hidrofóbicas orientadas em direção ao núcleo e suas bordas hidrofílicas, em direção ao lado externo. (b) Partícula LDL mostrando ApoB-100 mergulhada na capa externa da partícula.Parte (a) de Segrest, J. P., et al. Adv. Protein. Chem. 45: 303, Parte (b) de Schumaker, V. N., et al., Protein Chem. 45: 205,")
14
FIGURA 3.44Cadeias laterais de um segmento anfipático helicoidal de apoproteína C1 entre resíduos 32 e 53. A face polar mostra resíduos ácidos no centro e resíduos básicos na borda. No outro lado da hélice, resíduos hidrofóbicos formam uma face longitudinal não-polar.Redesenhado com permissão de Sparrow, J. T. e Gotto, A. M. Jr. CRC Crit. Rev. Biochem. 13:87, Direitos autorais (1983) da CRC Press, Inc., Boca Raton, FL.
 da CRC Press, Inc., Boca Raton, FL..")
16
FIGURA 3.46O sistema Chaperonina GroEL-GroES para dobramento de proteínas. (a) Modelo espaço preenchido do complexo chaperonina inteiro em uma vista de cima, olhando para baixo a partir da proteína de cap GroES. GroES liga-se ao lado cis de GroEL. GroES tem 7 subunidades polipeptídicas de peso molecular 10 kDa, e as estruturas cilíndricas cis e trans GroEL são compostas cada uma por 7 subunidades polipeptídicas (total 14), cada uma com 60 kDa. (b) Mesmo que (a), mas em vista lateral, com o anel trans GroEL em cinza-escuro e o cap GroES em cinza-claro. (c) Desenho de fi tas de um corte passando pelo centro do complexo chaperonina mostrando cavidades internas das secções cilíndricas cis e trans onde proteínas des-naturadas entram e ocorre seu dobramento. (d) Esquema simplifi cado de dobramento de proteína facilitado por GroEL. (i) A proteína desnaturada (linha curva preta) entra em GroEL e liga-se na cavidade com alta afi nidade. (ii) ATP liga-se a um anel produzindo uma conformação alterada. (iii) O anel ligado a ATP liga GroES para seqüestrar a proteína desnaturada na câmara de dobramento e uma grande mudança conformacional no anel ligado a GroES abre a proteína desnaturada para desfazer sua conformação não-produtiva e depois libera a proteína na cavidade expandida para dobrar espontaneamente. (iv) A proteína dobra-se dentro da câmara e ATP é hi-drolisado. (v) ATP liga-se ao anel trans, o que libera GroES e a proteína dobrada. (vi) Uma nova proteína é encapsulada. (e) Diagramas de energia livre para dobramento de proteínas. Esquerda: dobramento da proteína em ausência de chaperonina resulta em interme-diário retido cineticamente. Meio: no dobramento em chaperonina, empurrar e liberar (annealing) dissociam intermediários retidos e dá à proteína chance adicional de dobrar sem ser retida. Direita: além disso, a cavidade GroEL alisa o panorama energético para evitar a formação de certos intermediários retidos.Partes (a), (b) e (c) reproduzidas com permissão de Xu, Z. H., Horwicz, A. L. e Sigler, P. B. Nature 388:741, Direitos autorais (1997) Nature. Parte (d) reproduzida com permissão de Saibil, H. R. e Ranson, N. A. Trands Biochem. Sci. 27:627, © Elsevies. Ranson, N. A., et al., Cell 107:869, Parte (e) redesenhado de Ulrich, F., Hartl, F. e Hayer-Hartl, M. Science 295:1852, 2002.
 Modelo espaço preenchido do complexo chaperonina inteiro em uma vista de cima, olhando para baixo a partir da proteína de cap GroES. GroES liga-se ao lado cis de GroEL. GroES tem 7 subunidades polipeptídicas de peso molecular 10 kDa, e as estruturas cilíndricas cis e trans GroEL são compostas cada uma por 7 subunidades polipeptídicas (total 14), cada uma com 60 kDa. (b) Mesmo que (a), mas em vista lateral, com o anel trans GroEL em cinza-escuro e o cap GroES em cinza-claro. (c) Desenho de fi tas de um corte passando pelo centro do complexo chaperonina mostrando cavidades internas das secções cilíndricas cis e trans onde proteínas des-naturadas entram e ocorre seu dobramento. (d) Esquema simplifi cado de dobramento de proteína facilitado por GroEL. (i) A proteína desnaturada (linha curva preta) entra em GroEL e liga-se na cavidade com alta afi nidade. (ii) ATP liga-se a um anel produzindo uma conformação alterada. (iii) O anel ligado a ATP liga GroES para seqüestrar a proteína desnaturada na câmara de dobramento e uma grande mudança conformacional no anel ligado a GroES abre a proteína desnaturada para desfazer sua conformação não-produtiva e depois libera a proteína na cavidade expandida para dobrar espontaneamente. (iv) A proteína dobra-se dentro da câmara e ATP é hi-drolisado. (v) ATP liga-se ao anel trans, o que libera GroES e a proteína dobrada. (vi) Uma nova proteína é encapsulada. (e) Diagramas de energia livre para dobramento de proteínas. Esquerda: dobramento da proteína em ausência de chaperonina resulta em interme-diário retido cineticamente. Meio: no dobramento em chaperonina, empurrar e liberar (annealing) dissociam intermediários retidos e dá à proteína chance adicional de dobrar sem ser retida. Direita: além disso, a cavidade GroEL alisa o panorama energético para evitar a formação de certos intermediários retidos.Partes (a), (b) e (c) reproduzidas com permissão de Xu, Z. H., Horwicz, A. L. e Sigler, P. B. Nature 388:741, Direitos autorais (1997) Nature. Parte (d) reproduzida com permissão de Saibil, H. R. e Ranson, N. A. Trands Biochem. Sci. 27:627, © Elsevies. Ranson, N. A., et al., Cell 107:869, Parte (e) redesenhado de Ulrich, F., Hartl, F. e Hayer-Hartl, M. Science 295:1852,")
18
FIGURA 3.58Geração de fluxo eletro-osmótico em direção ao cátodo em eletroforese capilar.
FIGURA 3.61Cromatografia de exclusão molecular. Uma proteína pequena pode entrar nas partículas porosas de gel e será retardada na coluna em relação a uma proteína maior, que não pode entrar nas partículas porosas de gel.

19
FIGURA 3.63Resultado bidimensional (2-D) de proteínas expressas por células em cultura. (a) Proteínas solúveis de extrato celular (500 µg) aplicados ao gel e separados por focalização isoelétrica (pH 4,9- 5,5) na direção horizontal e por massa molecular na direção vertical (eletroforese em presença de detergente SDS). Mais de 1500 proteínas são observadas no gel por coloração com prata. (b) Uma região do gel expandida para mostrar detalhes. Proteí-nas numeradas foram analisadas por hidrólise com protease e espectroscopia de massa para determinar sua seqüência parcial de aminoácidos, levando à sua identificação.Reproduzido com premissão de Gygi, S. P., Corthals, G. L., Zhang, Y., Rochon, Y. e Aebersold, R. Proc. Natl Acad. Sci. USA 97:9390, 2000.
 Proteínas solúveis de extrato celular (500 µg) aplicados ao gel e separados por focalização isoelétrica (pH 4,9- 5,5) na direção horizontal e por massa molecular na direção vertical (eletroforese em presença de detergente SDS). Mais de 1500 proteínas são observadas no gel por coloração com prata. (b) Uma região do gel expandida para mostrar detalhes. Proteí-nas numeradas foram analisadas por hidrólise com protease e espectroscopia de massa para determinar sua seqüência parcial de aminoácidos, levando à sua identificação.Reproduzido com premissão de Gygi, S. P., Corthals, G. L., Zhang, Y., Rochon, Y. e Aebersold, R. Proc. Natl Acad. Sci. USA 97:9390,")
20
FIGURA 3.68Mapa de densidade eletrônica em resolução de 1,9-Å da região de sítio ativo da forma pró-enzima da tripsina. Resíduos de aminoácidos do sítio ativo são ajustados em um mapa de densidade eletrônica.Reimpresso com permissão de Kossiakoff, A. A., Chambers, J. L., Kay, L. M., e Stroud, R. M. Biochemistry 16:654, Direitos autorais (1977) da American Chemical Society FIGURA 3.69Tracejado estérico da estrutura do esqueleto de carbono-α da protease do HIV com inibidor (linhas grossas) e da protease sem inibidor (linhas finas).Redesenhado com permissão de Miller, M., Schneider, J., Sathyanarayana, B. K., Toth, M. V., Marshall, G. R., Clawson, L., Selk, L., Kent, S. B. H., e Wlodawer, A. Science 246:1149, Direitos autorais (1989) AAAS
 e da protease sem inibidor (linhas finas).Redesenhado com permissão de Miller, M., Schneider, J., Sathyanarayana, B. K., Toth, M. V., Marshall, G. R., Clawson, L., Selk, L., Kent, S. B. H., e Wlodawer, A. Science 246:1149, Direitos autorais (1989) AAAS.")
21
FIGURA 3.76Estrutura de NMR da proteína plastocianina de feijão francês. Estrutura mostra superposição de oito estruturas do esqueleto polipeptídeo da proteína, calculado a partir de restrições do espectro de NMR.De Moore, J. M., Lepre, C. A., Gippert, G. P., Chazin, W. J., Case, D. A. e Wright, P. E. J. Mol. Biol. 221: 533, Figura generosamente fornecida por P. E Wright

Apresentações semelhantes


 – Lápides 1, 2, 3» «nomes gravados, 21 de Agosto de 2008» «Ultramar.TerraWeb»>")




. Nenhuns direitos reservados, excepto para fins comerciais. Por favor, não coloque.>")








 forma não-ligada de hexoquinase e glicose livre e (b) conformação.>")




