Carregar apresentação
A apresentação está carregando. Por favor, espere

1
DNA mitocondrial na Antropologia
Ana von Hafe Pérez Manuel Lencastre Cardoso Maria Ramalho Fontes Pedro Leite Alves

2
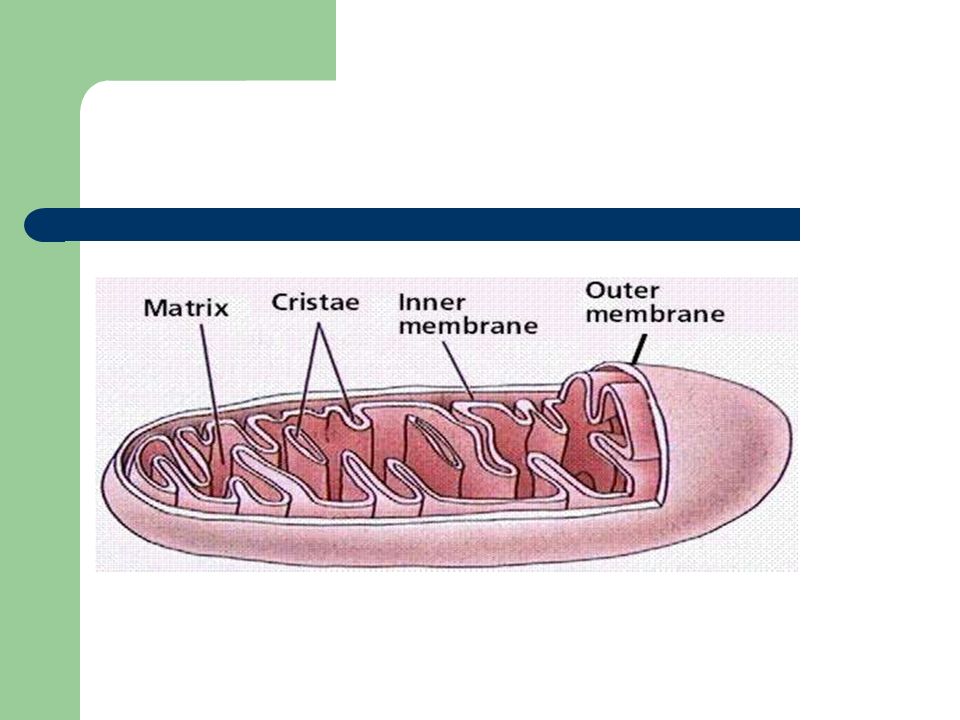
O que é a mitocôndria? É um organelo, notavelmente móvel e plástico, presente no citoplasma de células eucarióticas; É uma estrutura cilíndrica com, aproximadamente, µm de diâmetro; O conteúdo mitocondrial das nossas células é herdado apenas por via materna; É organizada em: matriz membrana interna membrana externa espaço intermembranar

4
Realiza a maior parte das oxidações celulares e produz a energia celular das células animais:
- O piruvato e os ác.gordos são convertidos em acetil CoA, o qual é oxidado a CO2 no ciclo de Krebs; - Grandes quantidades de NADH E FADH2 são produzidas por estas reacções de oxidação; - A energia disponível, pela combinação do oxigénio com os electrões levados pelo NADH e pelo FADH2, é regulada por uma cadeia transportadora de electrões na membrana mitocondrial interna, denominada cadeia respiratória.

6
Como terão surgido as mitocôndrias?
TEORIA AUTOGÉNICA A célula eucariótica teria surgido através da especialização de membranas internas, derivadas de invaginações da membrana plasmática.

7
TEORIA ENDOSSIMBIÓTICA Teoria proposta por Lynn Margulis.
A célula eucariótica seria o resultado da associação de células procarióticas simbióticas ( Bactéria Rickettsia prowazeki – Nature, 1998); A simbiose entre estas células procarióticas teria evoluído para graus de intimidade tais que algumas células envolveriam outras completamente, embora as 1as ficassem intactas no interior do hospedeiro;
; A simbiose entre estas células procarióticas teria evoluído para graus de intimidade tais que algumas células envolveriam outras completamente, embora as 1as ficassem intactas no interior do hospedeiro;")
9
Factos que apoiam a teoria endossimbiótica
Material genético igual entre procariontes e eucariontes; Simbiose é um processo muito comum no mundo vivo; Tamanho dos cloroplastos e mitocôndrias muito semelhante ao dos procariontes actuais; Síntese proteica das mitocôndrias e cloroplastos é inibida por substâncias inibidoras de procariontes (estreptomicina e cloranfenicol) mas não por inibidores de eucariontes ( cicloheximida); A.a iniciador da cadeia polipeptídica de uma mitocôndria ou cloroplasto é a formil-metionina, como nas bactérias e não a metionina, como nos eucariontes.
 mas não por inibidores de eucariontes ( cicloheximida); A.a iniciador da cadeia polipeptídica de uma mitocôndria ou cloroplasto é a formil-metionina, como nas bactérias e não a metionina, como nos eucariontes.")
10
O que é o DNA mitocondrial?
O DNAmt humano contém pares de bases - um dos mais pequenos que se conhece. Constituído, normalmente, por uma molécula circular de DNA em cadeia dupla; Localizado na matriz, por vezes ligado à membrana interna; Todos os seus transcritos e os seus produtos de tradução ficam no organelo – não há exportação de RNA ou proteínas.

11
O DNAmt codifica: - 2 RNA ribossómicos - 22 RNA de transferência - 13 sequências codificantes de proteínas Todos estes produtos de tradução foram identificados: - 3 são subunidades do complexo cit c oxidase - 2 são subunidades da ATPase - 7 são subunidades do complexo NADH – CoQ reductase - 1 (cit b) é a subunidade do complexo CoQ cit c reductase
 é a subunidade do complexo CoQ cit c reductase.")
12
Como terá evoluído o DNAmt?
Durante a evolução inicial das células eucarióticas, ocorreu, provavelmente, uma transferência maciça de genes do organelo (mitocôndria) para o núcleo; É possível que esta transferência tenha cessado antes de estar concluída; Restaram ainda alguns genes na mitocôndria ( os outros passaram para o núcleo) – DNA mitocondrial
 para o núcleo; É possível que esta transferência tenha cessado antes de estar concluída; Restaram ainda alguns genes na mitocôndria ( os outros passaram para o núcleo) – DNA mitocondrial.")
13
Dados que apoiam a Antropologia
Paleontológicos - Fósseis Genéticos – DNA mitocondrial Cromossoma Y DNA nuclear

14
Dados Paleontológicos

15
Porquê o DNA mitocondrial?
Cada célula humana tem um “segundo” genoma encontrado na mitocôndria. Isto quererá dizer que o genoma mitocondrial está altamente amplificado. Enquanto cada célula contém apenas duas cópias de um dado gene nuclear (1 de cada par de homólogos), há milhares de cópias de um dado gene mitocondrial por célula. Assim o DNA mitocondrial é o sistema genético adoptado em casos onde o tecido da amostra estão muito velhos, são muito pequenos ou foram degradados pelo calor ou humidade.
, há milhares de cópias de um dado gene mitocondrial por célula. Assim o DNA mitocondrial é o sistema genético adoptado em casos onde o tecido da amostra estão muito velhos, são muito pequenos ou foram degradados pelo calor ou humidade.")
16
Porquê o DNA mitocondrial?
O DNA mitocondrial é o DNA humano mais fácil de amplificar através de PCR. Este método examina uma sequência de 440 nucleotídeos de uma região não codificada do genoma mitocondrial. O DNA mitocondrial não recombina. Há assim uma história genealógica imutável. DNA mitocondrial é herdado somente da mãe.

17
Herança materna do DNA mitocondrial
A mitocôndria e o seu genoma são herdados da linha materna. As crianças do sexo masculino partilham o genoma mitocondrial com a sua mãe e parentes femininos. Os pais não têm qualquer contribuição para o património mitocondrial dos filhos. O genoma mitocondrial é herdado intacto através de milhares de gerações. Embora algumas novas mutações apareçam ocasionalmente numa linhagem são muito poucas e fáceis de identificar.

18
Herança materna do DNA mitocondrial

19
Análise ao cromossoma Y
É o segundo sistema genético mais utilizado no estudo da população humana. Herdado pela linha paterna e assim complementa o mtDNA. 97% não recombina (só recombinam as zonas I e II).
.")
20
DNA nuclear Maior amplitude genética.
No entanto sofre recombinação e possui mecanismos de correcção mutacional.

21
“Where do we come from?”

22
Homo Erectus Todos os cientistas estão de acordo que há cerca de 1,7 milhão de anos houve uma migração de uma espécie de hominídios anterior ao Homo Sapiens chamada Homo Erectus que a partir de África colonizou os outros continentes.

23
Há 100 000 anos havia no mundo vários hominídios diferentes:
-Em África e no médio oriente » Homo Sapiens -Na Ásia » Homo Erectus -Na Europa » Homo Neanderthalensis Há anos toda essa diversidade tinha já desaparecido e todas as populações eram então constituídas por seres humanos anatomicamente e comportamentalmente modernos.

24
Modelos Modelo Multiregional ou de Continuidade “Out of Africa” ou Modelo de Substituição

25
Modelo Multiregional As primeiras referências a este modelo foram feitas por Franz Weidenreich em 1943 mas ele foi largamente desenvolvido por Milford Wolpoff e Alan Thorne Esta modelo defende que depois da dispersão do Homo Erectus se manteve um fluxo de genes suficiente para que surgisse o Homo Sapiens de forma lenta e progressiva fruto das várias migrações entre as populações de todo o mundo habitado, logo os Homo Neanderthalensis não eram uma espécie mas uma população que contribuiu geneticamente para a formação do homo sapiens; Esse fluxo génico não permitiu a especiação das populações dispersas (Wolpoff and Caspari,1997); As variações regionais são fruto da selecção natural desde a dispersão do Homo Erectus; Esta teoria baseia-se essencialmente em argumentos paleontológicos, sobretudo na continuidade encontrada em caracteres de vários fosseis.
; As variações regionais são fruto da selecção natural desde a dispersão do Homo Erectus; Esta teoria baseia-se essencialmente em argumentos paleontológicos, sobretudo na continuidade encontrada em caracteres de vários fosseis.")
26
Alan Templeton e o modelo multiregional
Colectou todos os trabalhos previamente feitos acerca do tema e tratou-os com métodos diferentes (Programa de análise estatística GEODIS) Deixou de dar tanto ênfase às variações da ansa D do mtDNA, dando-se mais importância a outras zonas de sinal filogenético mais antigo); Considerou no seu estudo: Cromossoma Y; 2 regiões do cromossoma X; 6 regiões autossómicas Templeton concluiu que: Existe de facto um papel dominante de África na modelagem de todo o património genético através de pelo menos 2 expansões a partir do grupo inicial de Homo Erectus; Há ocorrência de fluxo genético entre populações que levou a intercruzamentos e não a substituição (Cann et al., 1987)
 Deixou de dar tanto ênfase às variações da ansa D do mtDNA, dando-se mais importância a outras zonas de sinal filogenético mais antigo); Considerou no seu estudo: Cromossoma Y; 2 regiões do cromossoma X; 6 regiões autossómicas. Templeton concluiu que: Existe de facto um papel dominante de África na modelagem de todo o património genético através de pelo menos 2 expansões a partir do grupo inicial de Homo Erectus; Há ocorrência de fluxo genético entre populações que levou a intercruzamentos e não a substituição (Cann et al., 1987)")
27
“Out of Africa” Em 1987 na revista nature Rebecca Cann publicou um artigo com um modelo novo baseado em argumentos essencialmente genéticos; Eles defendem que depois da migração inicial as populações de Homo Erectus dispersas deram origem por radiação adaptativa a várias espécies; o Homo Sapiens, surgido em África há ou anos, substituiu as populações existentes em todo o mundo sem trocas significativas de genes; As variações regionais são um fenómeno muito recente; os autores desprezam os registos fosseis afirmando que não são suficientemente numerosos para serem estatisticamente válidos.

28
“Mitochondrial DNA and human evolution” by Rebecca L
“Mitochondrial DNA and human evolution” by Rebecca L. Cann, Mark Stoneking and Allan C. Wilson Foram utilizados no estudo 145 amostras placentárias e duas linhas celulares segundo as seguintes regiões: África sub-Sahárica (20); Ásia (34); Europa, Norte de África e Médio Oriente (46); Austrália (21); Papua Nova Guiné (26) os autores fizeram mapas de restrição dos DNAmt das amostras especialmente na zona da ansa D (D loop);
; Ásia (34); Europa, Norte de África e Médio Oriente (46); Austrália (21); Papua Nova Guiné (26) os autores fizeram mapas de restrição dos DNAmt das amostras especialmente na zona da ansa D (D loop);")
29
“Mitochondrial DNA and human evolution” by Rebecca L
“Mitochondrial DNA and human evolution” by Rebecca L. Cann, Mark Stoneking and Allan C. Wilson As conclusões deste estudo apresentam-se sob a forma de um esquema que traduz uma possível origem comum a todos os seres humanos modernos : Existem então duas ramificações primárias; uma composta inteiramente de Africanos e outra incluindo todas as outras populações; Cada população tem linhagens múltiplas que contactam com a árvore em diferentes posições; Na elaboração da Árvore de máxima parcimónia assumiram-se duas condições: Um modo de transmissão inteiramente maternal de DNA mitocondrial; Cada indivíduo possui um conjunto homogéneo de múltiplos genomas mitocôndriais; Conclui-se desta forma que a origem dos modernos humanos se deu em África à entre de há a anos atrás;

30
“Global Patterns of Linkage Desiquilibrium at the CD4 Locus and Modern Human Origins” Tishkoff et al,1996 Um estudo internacional recolheu 1600 amostras de DNA cromossómico de 42 regiões geograficamente distintas. Da análise ao locus CD4 do cromossoma 12 concluiu-se que todas as populações excepto as sub-Saharianas apresentam delecção do ALU e um único tipo de STRP (Small Tandem Repeats Polymorphism). (Tishkoff et al,1996)
. (Tishkoff et al,1996)")
31
“Out of africa” vs. Multiregionalismo
Qual a mais correcta?

32
“Out of Africa” – Argumentos favoráveis
→ Muito do suporte desta teoria advém de estudos genéticos nomeadamente do estudo do DNAmt A análise das mutações neutras sofridas pelo DNAmt, nomeadamente na região da D loop foi usada para determinar uma suposta origem de toda a linhagem humana moderna (Cann et al,1987)
")
33
“Out of Africa” – Argumentos favoráveis
O estudo de DNA nuclear de 5 populações geograficamente separadas actualmente permitiu chegar à conclusão de que existe maior variabilidade genética no continente africano do que nos restantes o que pode indiciar uma origem neste continente de toda a linhagem humana moderna (Gibbons et al,1992;)
")
34
“Out of Africa” – Argumentos favoráveis
Vários estudos apontam para uma transição de humanos arcaicos para humanos modernos há cerca de anos atrás: Estudos comparativos de polimorfismos em 30 microsatélites cromossómicos levaram à conclusão da provável existência de uma população ancestral africana nessa altura (Goldstein et al,1995) Um estudo genético analisou exaustivamente todas as bp do DNAmt de três humanos (1 africano, 1 europeu e 1 asiático) e 4 especímenes símios, chegando à conclusão que a separação humana dos macacos ocorreu há 13 milhões de anos e a divisão da população africana e não-africana deu-se há ± anos (Horai et al, 1995) Análises a fósseis de humanos modernos datados de há anos encontrados na Etiópia e África do Sul (Gore et al,1997) A determinação do relógio molecular do DNA mitocondrial, ou seja, a taxa à qual ocorreram mutações no DNAmt ao longo do tempo (Nei et al,1995)
 Um estudo genético analisou exaustivamente todas as bp do DNAmt de três humanos (1 africano, 1 europeu e 1 asiático) e 4 especímenes símios, chegando à conclusão que a separação humana dos macacos ocorreu há 13 milhões de anos e a divisão da população africana e não-africana deu-se há ± anos (Horai et al, 1995) Análises a fósseis de humanos modernos datados de há anos encontrados na Etiópia e África do Sul (Gore et al,1997) A determinação do relógio molecular do DNA mitocondrial, ou seja, a taxa à qual ocorreram mutações no DNAmt ao longo do tempo (Nei et al,1995)")
35
“Out of Africa” – Argumentos favoráveis
Alguns cientistas afirmam que a população ancestral de Homo Sapiens era diminuta, na ordem dos indivíduos (Rogers and Harpending, 1992; Harpending et al, 1993; Nei and Takahata, 1993; Harpending, 1994) Novas espécies costumam surgir de pequenas e isoladas populações, uma vez que o isolamento mantém e acentua a diversidade genética das populações, podendo conduzir à especiação.
 Novas espécies costumam surgir de pequenas e isoladas populações, uma vez que o isolamento mantém e acentua a diversidade genética das populações, podendo conduzir à especiação.")
36
“Out of Africa” – Argumentos desfavoráveis
Crânios fósseis com características mistas de Homo Erectus e Homo Sapiens foram encontrados no Sudeste Asiático indiciando um possível intercruzamento (Wolpoff et al, 1992) A descoberta recente em Portugal de um fóssil que reúne caracteres de Neandertais e de Homo Sapiens veio abalar a crença na substituição das populações de hominídeos na Europa (Duarte et al,1999) A descoberta de um fóssil no continente australiano que combina caracteres de Homo Sapiens e de Homo Erectus nativos da ilha é mais um argumento paleontológico que vem confirmar a ocorrência de fluxo genético entre as duas populações
 A descoberta recente em Portugal de um fóssil que reúne caracteres de Neandertais e de Homo Sapiens veio abalar a crença na substituição das populações de hominídeos na Europa (Duarte et al,1999) A descoberta de um fóssil no continente australiano que combina caracteres de Homo Sapiens e de Homo Erectus nativos da ilha é mais um argumento paleontológico que vem confirmar a ocorrência de fluxo genético entre as duas populações.")
37
“Out of Africa” – Argumentos desfavoráveis
Não é cientificamente comprovável a existência de superioridade intelectual e comportamental de Homo Sapiens sobre Homo Erectus (Wolpoff et al,1992) O facto da variação genética ser maior no continente africano pode ser um indicativo de um maior número de habitantes em comparação com as outras regiões ao invés de maior antiguidade (Relethford, 2001) Os dados utilizados por R. L. Cann e colaboradores para a elaboração de uma árvore de máxima parcimónia foram analisados por outros cientistas que chegaram a diversas árvores revelando-se este método altamente subjectivo (Templeton, 2002)
 O facto da variação genética ser maior no continente africano pode ser um indicativo de um maior número de habitantes em comparação com as outras regiões ao invés de maior antiguidade (Relethford, 2001) Os dados utilizados por R. L. Cann e colaboradores para a elaboração de uma árvore de máxima parcimónia foram analisados por outros cientistas que chegaram a diversas árvores revelando-se este método altamente subjectivo (Templeton, 2002)")
38
“Out of Africa” – Argumentos desfavoráveis
O ênfase na região D-loop veio a provar-se errónea uma vez que a taxa de mutação é de tal maneira elevada que não apresenta valor evolutivo para a determinação de uma história passada mas sim de afinidades recentes: “Back mutation”; locais em que ocorreram sucessivas mutações que conduziram à sequência inicial de bases “Parallel substitution”; mutações ocorrem no mesmo local para linhagens diferentes “Rate heterogeneity”; dentro da mesma região existem locais com diferentes susceptibilidades a sofrerem mutações.

39
“Out of Africa” – Argumentos desfavoráveis
Análises feitas ao gene PDHA1 do cromossoma X através da sua taxa e velocidade mutacional determinaram a existência de duas linhagens evolutivas de Homo Sapiens ao invés de uma única (Hey et al,1999) A existência de mutações no DNAmt pode ter levado à acção da selecção natural sobre os indivíduos seus portadores, desafiando dessa forma a validade do relógio molecular.
 A existência de mutações no DNAmt pode ter levado à acção da selecção natural sobre os indivíduos seus portadores, desafiando dessa forma a validade do relógio molecular.")
40
“Out of Africa” – Argumentos desfavoráveis
Estimativas da altura da coalescência baseadas em análises de genes humanos de β-globina apontam para a data de há anos atrás, contrariando assim a teoria da origem recente do Homem (Fullerton et al,1997)
")
41
Teoria Multiregional – Argumentos favoráveis
→ Utiliza essencialmente dados paleontológicos na sua fundamentação Descobertas de crânios com características mistas de Homo Erectus e Homo Sapiens no sudeste asiático datados de há anos representam uma das maiores provas de que existiu fluxo genético entre estes dois grupos (Macintosh et al, 1974; Wolpoff et al, 1984) A recente descoberta de um fóssil de uma criança de 4 anos que reúne características de Homo Neandertalensis e Homo Sapiens em Portugal, no Abrigo do Lagar Velho datada de há anos está a gerar grande controvérsia na comunidade científica defensora da teoria da substituição sendo até considerada como o argumento mais credível para a existência de fluxo genético (Trinkaus et al,1999)
 A recente descoberta de um fóssil de uma criança de 4 anos que reúne características de Homo Neandertalensis e Homo Sapiens em Portugal, no Abrigo do Lagar Velho datada de há anos está a gerar grande controvérsia na comunidade científica defensora da teoria da substituição sendo até considerada como o argumento mais credível para a existência de fluxo genético (Trinkaus et al,1999)")
42
Teoria Multiregional – Argumentos favoráveis
A ocorrência de uma mutação de um gene específico para a anemia falciforme ocorrida na Europa anteriormente à migração africana que se manteve até à actualidade indicia que terá havido fluxo genético entre as populações ancestrais europeias e as modernas africanas (Templeton, 2002)
")
43
Teoria Multiregional – Argumentos favoráveis
A descoberta da “Eva mitocondrial” é interpretada pelos multiregionalistas como um episódio relativamente frequente na altura e que se difundiu pelo Velho Mundo não por competição mas por fluxo genético; Através de estudos cromossómicos, foi possível determinar que ocorreram no mínimo 2 grandes migrações para fora de África depois da migração original de Homo Erectus (Templeton,2002): Há anos (DNAmt e Cromossoma Y); Há anos (Regiões autossomais e “X-linked”)
: Há anos (DNAmt e Cromossoma Y); Há anos (Regiões autossomais e X-linked )")
44
Teoria Multiregional – Argumentos favoráveis
Uma nova análise de DNA nuclear está a pôr em causa a ideia de que uma pequena população de ancestrais africanos deu origem a toda a variedade actual. O estudo de uma região específica do cromossoma X (PDHA1) em grande variedade populacional actual levou à conclusão da migração de duas pequenas populações de Homo Sapiens para locais geográficos diferentes do Velho Mundo ocorrendo co-evolução através de fluxo genético entre esses dois grupos e os ancestrais de cada zona (Hey and Harris,1999)
 em grande variedade populacional actual levou à conclusão da migração de duas pequenas populações de Homo Sapiens para locais geográficos diferentes do Velho Mundo ocorrendo co-evolução através de fluxo genético entre esses dois grupos e os ancestrais de cada zona (Hey and Harris,1999)")
45
Teoria Multiregional – Argumentos desfavoráveis
O registo fóssil é uma base argumentativa com algumas falhas: Raros exemplares; Fósseis em estado degradado; Dificuldades de datação; Não representam toda a variedade fenotípica da sua época; O seu tratamento pode influenciar as suas informações. O facto de ter ocorrido evolução paralela de diferentes raças que acumularam certos traços distintos conforme o meio ambiente em que vivem pode levar a interpretações racistas desta teoria, como aconteceu na sua génese no período da Alemanha Nazi

46
Teoria Multiregional – Argumentos desfavoráveis
A evolução paralela ocorrida em locais com diferentes pressões ambientais como é defendida pelos multiregionalistas é criticada pelos biólogos uma vez que colide com os princípios de Darwin É altamente improvável a não-ocorrência na evolução humana de competição inter-específica com sobreposição de uma espécie e consequente eliminação de outras, uma vez que este fenómeno ocorre a todos os níveis da Natureza

47
“Out of Africa” CONCLUSÃO
O modelo mais aceite actualmente pela comunidade científica é: “Out of Africa”

48
Criticas aos métodos Muitos dos argumentos que suportam quer uma teoria quer outra são altamente subjectivos e falíveis pois os métodos utilizados na sua análise encontram-se pouco desenvolvidos Deste modo surgem agora novos avanços técnicos: Métodos estatísticos fiáveis (Programa GEODIS,…) Obtenção de amostras não contaminadas Análises genéticas mais abrangentes e mais reprodutíveis
 Obtenção de amostras não contaminadas. Análises genéticas mais abrangentes e mais reprodutíveis.")
49
Terceiro modelo Surgiu recentemente uma hipótese de terceiro modelo explicativo que se baseia nas duas hipóteses anteriores; Este terceiro modelo proposto por Triesman postula que: Ocorreu uma mutação há anos numa população africana isolada originando um novo genoma mitocondrial; Essa mutação tornou-se dominante ao mesmo tempo que essa população se expandiu para fora de África; Por migrações sucessivas (com fluxos genéticos significativos) esse genótipo mitocondrial tornou-se dominante nas várias populações do velho mundo;
 esse genótipo mitocondrial tornou-se dominante nas várias populações do velho mundo;")
50
Criticas a Triesman Não conseguimos encontrar nenhum artigo cientifico que rebatesse a teoria de Triesman; Encontramos no entanto vários comentários de notáveis cientistas conotados com um ou outro modelo pré-existente que negam a possibilidade de se ter passado qualquer outro cenário que não o descrito por esses dois modelos…

51
“Com os dados actuais, qualquer biólogo que estivesse e estudar a filogenia da drosophilla não hesitaria sobre o modelo a aplicar, mas quando o estudo incide sobre a nossa própria filogenia não somos capazes de afastar completamente as nossas tendências e emoções mais profundas…” Tattersaull

Apresentações semelhantes

















 teríam surgido das modificações genéticas que ocorrem com o.>")


