Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Controle e Mecanismos de Divisão
Ciclo celular: Controle e Mecanismos de Divisão Profa. Dra. Lucilene Regina Maschio

2
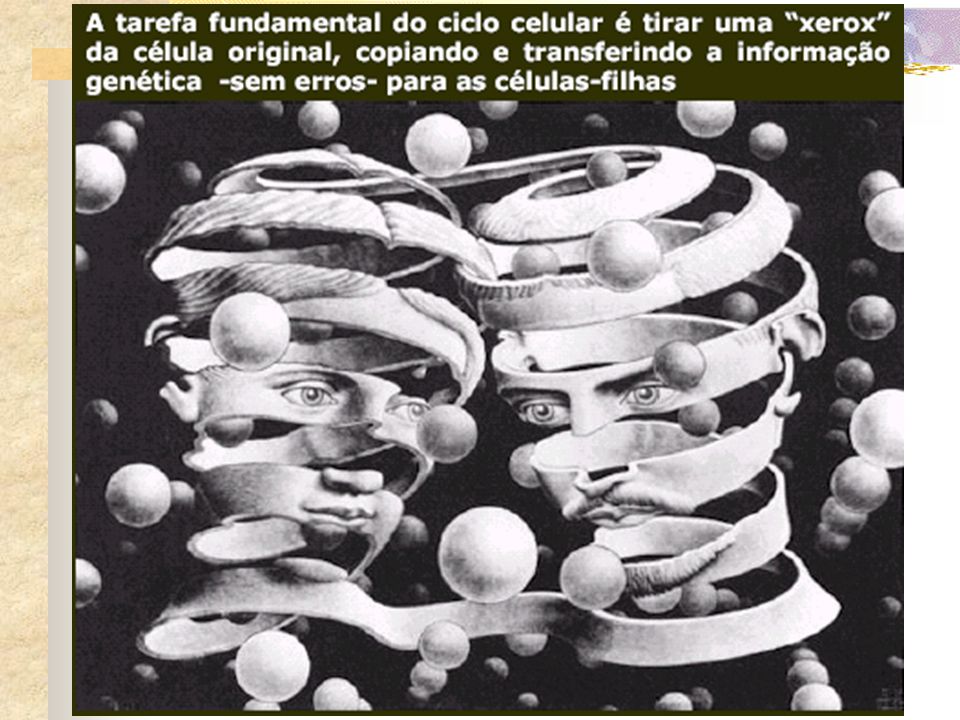
É a divisão de uma célula
Um dos fenômenos mais intrigantes de toda natureza Duplicam perpetuando a vida Se houver caminho para imortalidade Este encontra-se nos mecanismos que regem esta duplicação

3
POR QUE UMA CÉLULA SE DIVIDE?
Reprodução de organismos unicelulares e multicelulares Crescimento dos indivíduos: embrião adulto Reposição de células mortas ou lesadas: tecido epitelial, hepático e sangüíneo

4
Todas as células de um indivíduo adulto contêm a mesma informação genética.
mecanismo Primeiro replica o material genético e depois divide uma cópia completa da informação genética para cada célula-filha. Esse ciclo celular compreende, portanto, uma série de eventos pelos quais as células progride de uma divisão para a próxima.

5
VISÃO GERAL DO CICLO CELULAR

7
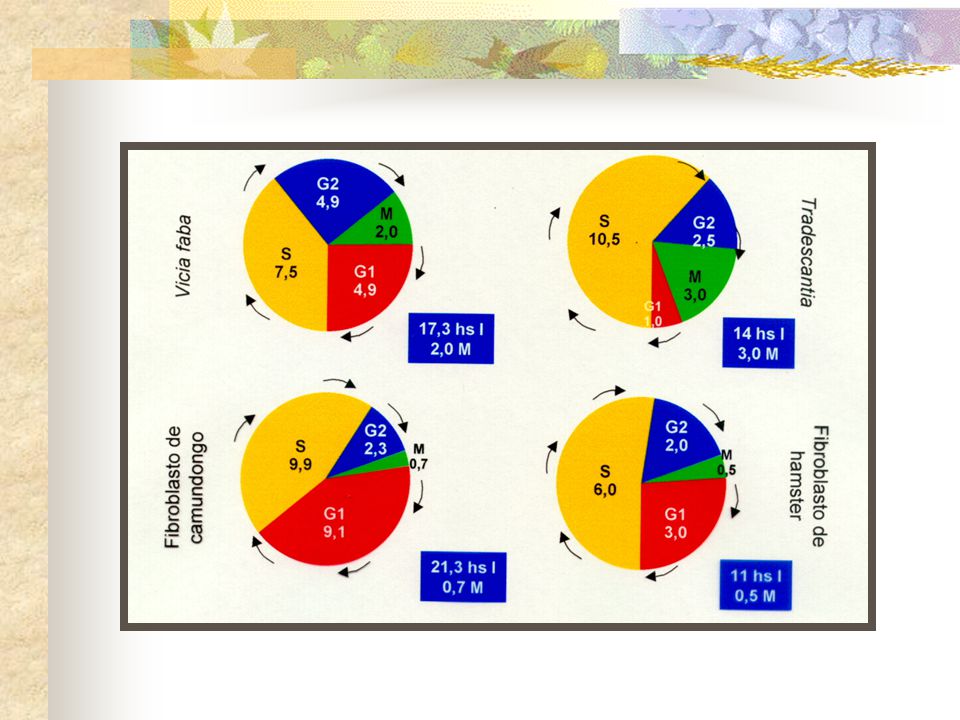
A duração do ciclo celular varia muito de um tipo de célula para outro.

8
CICLO CELULAR EM EMBRIÃO
. Os estágios iniciais de desenvolvimento de células embrionárias são caracterizados por divisões rápidas e sincronizadas. . As primeiras poucas divisões celulares após a fertilização servem para subdividir uma célula-ovo gigante em várias células menores, o quão rápido possível.

11
Para explicar como as células se reproduzem devemos considerar três questões principais:
1. Como as células duplicam o seu conteúdo? 2. Como elas repartem o conteúdo duplicado e se separam em duas? 3. Como elas coordenam toda a maquinaria que é necessária para esses dois processos?

12
CICLO CELULAR DE EUCARIOTOS

13
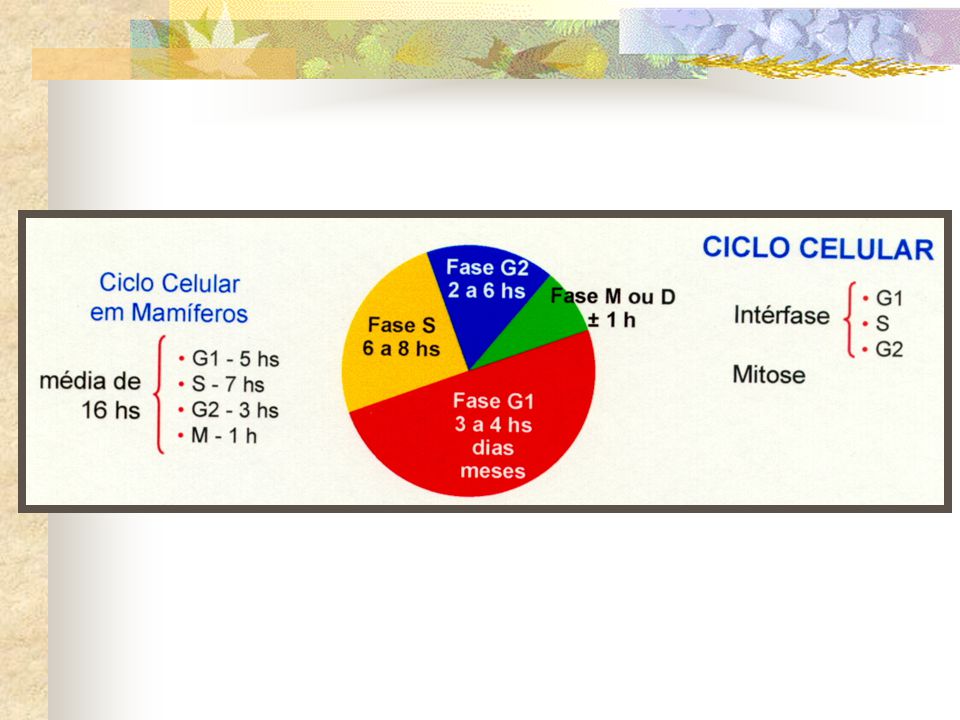
CICLO CELULAR 4 fases

14
1. Como as células duplicam o seu conteúdo?

15
INTÉRFASE No período entre duas mitoses, denominado intérfase, os cromossomos ocorrem como filamentos distendidos e espalhados por toda área nuclear, não sendo individualmente visualizados ao microscópio de luz. . A intérfase é um período de intensa síntese de todos os constituintes celulares. . Quando a célula recebe o sinal para divisão, sintetiza os componentes necessários para esse processo e dobra seu volume para possibilitar que duas novas células aproximadamente iguais no tamanho e composição sejam originadas por mitose.

16
Período G1 (G, do inglês, gap = intervalo)
Célula permanece em intérfase por anos sem se dividir . A subfase G1 é a que tem o tempo de duração mais variável. . Em muitas células eucarióticas, G1 dura de três a quatro horas, mas pode prolongar-se por dias meses ou anos, de acordo com as condições fisiológicas.

17
Neurônios: dependendo da lesão podem voltar ao ciclo celular
Algumas como linfócitos do sangue humano:presença de um antígeno e hepatócitos: perda do tecido hepático podem voltar a se dividir se houver um estímulo. Células de mamíferos privadas de fatores de crescimento interrompem o ciclo celular durante G1 num estágio chamado G0

18
Nesse período ocorre também:
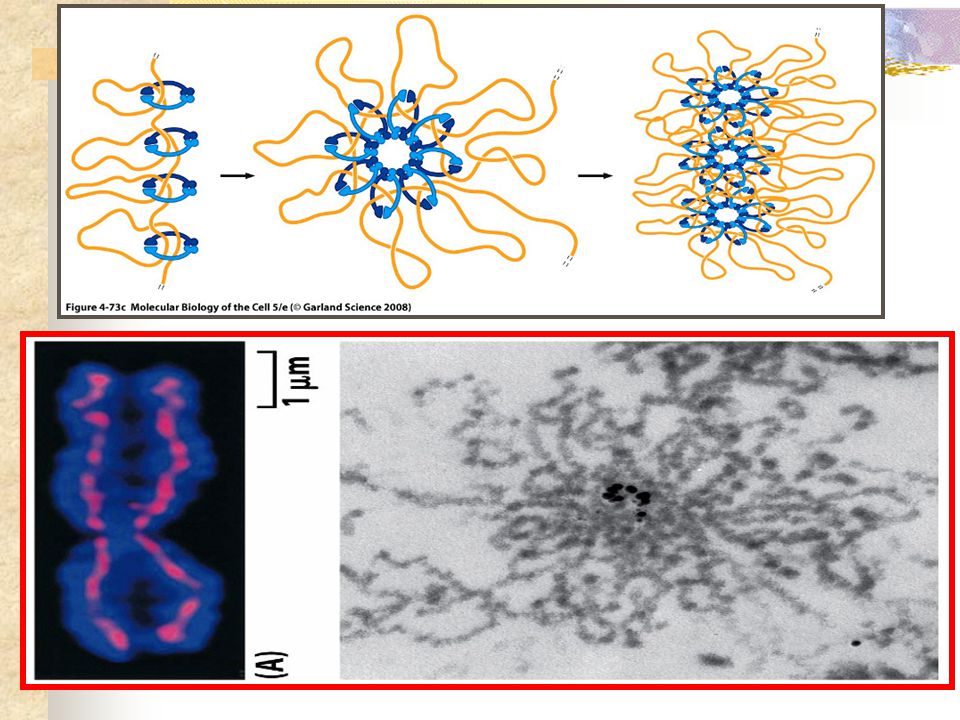
. Máxima síntese de proteínas não histonas; . Intensa síntese de proteínas no citoplasma; . Intensa síntese de RNAm, RNAt e RNAr; . Síntese de proteínas responsáveis pela replicação de DNA e duplicação dos centrossomos - CdKs . Síntese do complexo de proteínas COESINAS; . Síntese do complexo protéico CONDENSINA

19
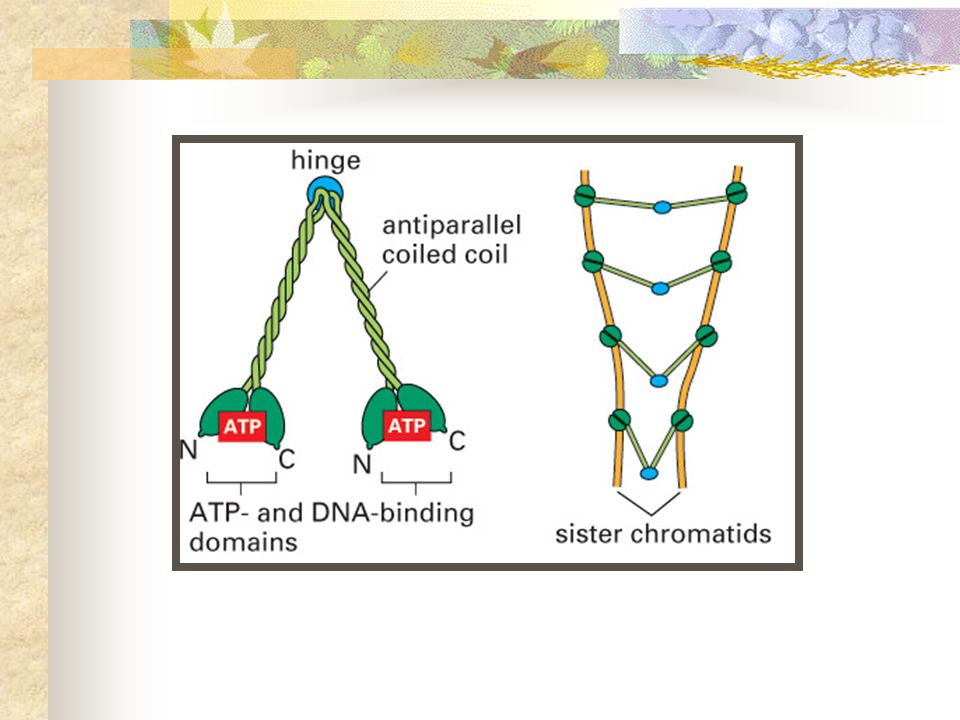
COESINAS S G1 4 subunidades: 1 heterodímero de Smc1 e Smc3 ( Structural maintenance of chromosome), que se associam a Scc1 e esta a Scc3 (Sister chromatid cohesion) . Heterodímero forma dois longos braços em espiral-espirada antiparalela, cujas cabeças (extremidades N e C-terminais_ têm função ATPásica.
, que se associam a Scc1 e esta a Scc3 (Sister chromatid cohesion) . Heterodímero forma dois longos braços em espiral-espirada antiparalela, cujas cabeças (extremidades N e C-terminais_ têm função ATPásica.")
21
CONDENSINAS

23
Período S 1. Duplicação do centrossomo;
2. Gatilho para replicação do DNA (mecanismo do tudo ou nada); 3. Síntese de histonas.
; 3. Síntese de histonas.")
24
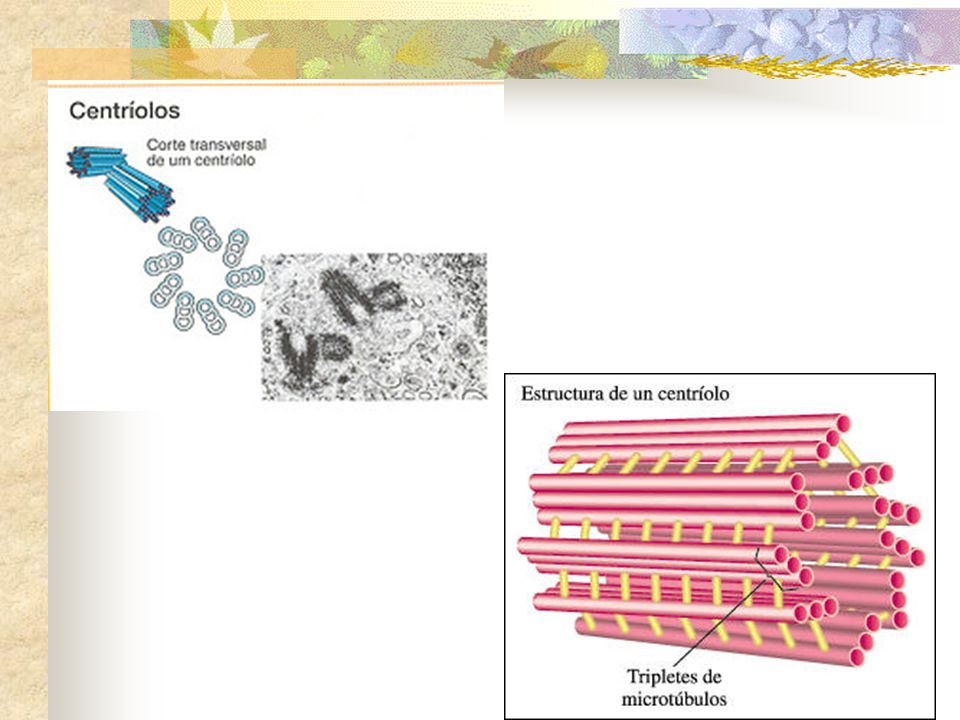
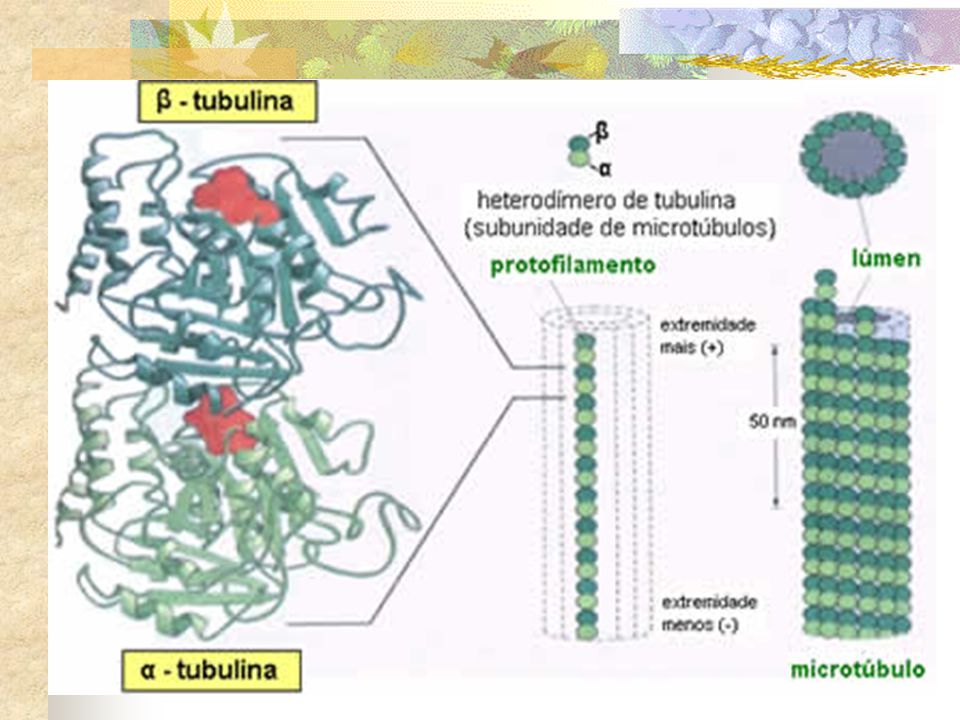
1.CENTROSSOMOS Localizado próximo ao núcleo da célula em intérfase e contém, na maioria das células animais, um par de centríolos. Centrossomo Microtúbulos Envoltório nuclear

27
Fases do ciclo do centrossomo:
Duplicação semi-conservativa: inicia no final de G1 e início da fase S Maturação do centrossomo: ocorre em G2 Separação dos centrossomos: transição G2-M Desorientação dos centrossomos: final da mitose e início de G1

29
Célula de mamífero na fase S, após a duplicação dos centríolos

30
FUNGOS C. VEGETAIS O. HUMANOS
. Em fungos, na maioria das plantas e nos ovócitos humanos, os centros organizadores de microtúbulos não contêm centríolos. FUNGOS microtúbulos são nucleados a partir de uma estrutura denominada corpúsculo polar do fuso, que se encontra embebido no envoltório nuclear Microtúbulos emanam de centros organizadores de microtúbulos distribuídos ao redor do envoltório nuclear C. VEGETAIS O. HUMANOS Um centro organizador acentriolar é responsável pela nucleação dos microtúbulos. Apesar da ausência dos centríolos, o fuso de divisão celular se forma normalmente uma vez que todas essas células contêm -tubulina, que é necessária para a nucleação dos microtúbulos.

31
2. REPLICAÇÃO DO DNA Garante que as células-filhas possam receber uma cópia exata de cada molécula de DNA da célula parental.

32
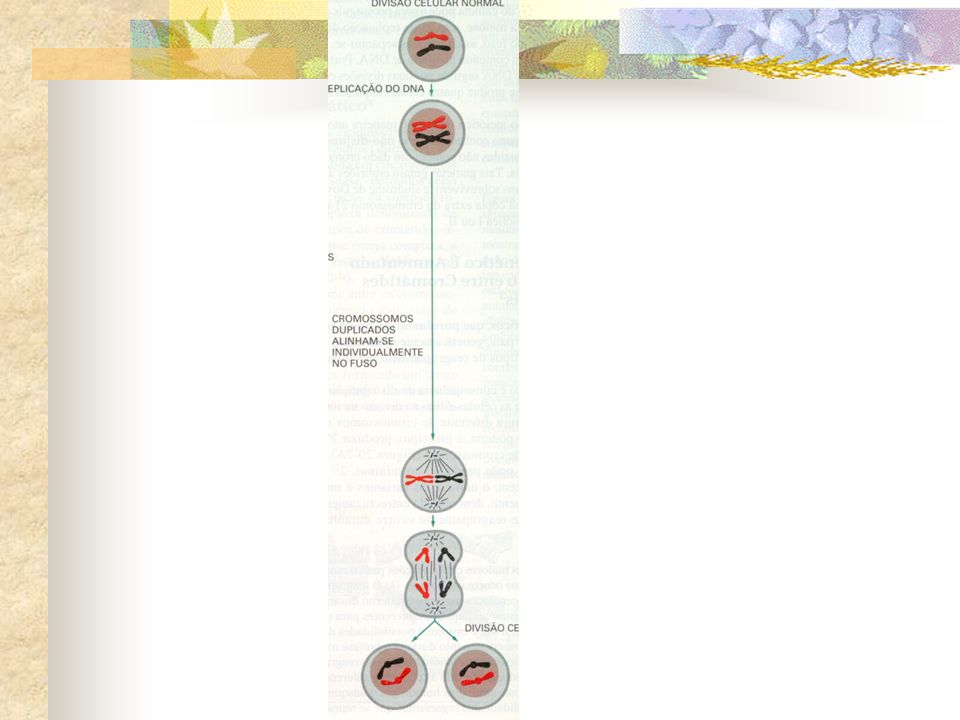
Células humanas diplóides Têm 2n = 46 cromossomos Célula em G1 Essas células continuam diplóides, contendo 2n = 46 cromossomos, embora com o dobro do conteúdo de DNA (4C) 46 moléculas de DNA (uma molécula para cada um dos 23 pares de homólogos) Em G2, a célula humana contém 92 moléculas de DNA, sendo que cada um dos 46 cromossomos contém duas moléculas de DNA (denominadas cromátidas irmãs) Durante a fase S, cada molécula de DNA dá origem a outra idêntica a ela.
 46 moléculas de DNA (uma molécula para cada um dos 23 pares de homólogos) Em G2, a célula humana contém 92 moléculas de DNA, sendo que cada um dos 46 cromossomos contém duas moléculas de DNA (denominadas cromátidas irmãs) Durante a fase S, cada molécula de DNA dá origem a outra idêntica a ela.")
33
2. SÍNTESE DE HISTONAS

34
Período G2 . Período em que a célula verifica, por exemplo, se todo DNA duplicou corretamente e se houve aumento adequado do volume, antes de iniciar a divisão celular propriamente dita. . Portanto as subfases S e G2, ocorrem somente em células que irão se dividir.

35
2. Como elas repartem o conteúdo duplicado e se separam em duas?

36
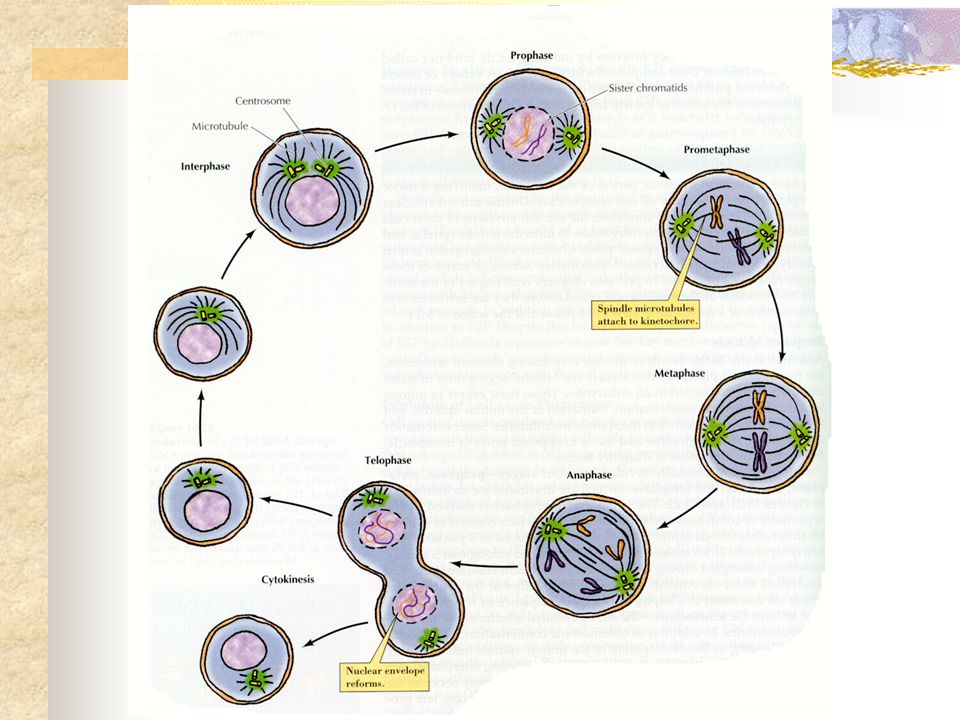
MITOSE Fase final de um processo que teve início em nível molecular com a duplicação do DNA . O processo de divisão celular é essencialmente o mesmo em todos os eucariotos, funcionando de maneira altamente controlada, de forma a garantir que, a cada ciclo de divisão, duas células idênticas sejam originadas.

37
MITOSE Condensação cromossômica Formação do fuso mitótico
Desorganização do envelope nuclear Ligação dos microtúbulos do fuso aos cinetócoros dos cromossomos Migração das cromátides-irmãs para pólos opostos Organização do envelope nuclear Descondensação dos cromossomos Citocinese: divide a célula

39
FASES DA MITOSE 1º) Prófase 2º) Pro-Metáfase 3º) Metáfase 4º) Anáfase 5º) Telófase
 Prófase 2º) Pro-Metáfase 3º) Metáfase 4º) Anáfase 5º) Telófase")
40
Um centrossomo inicia a formação do fuso mitótico
Caracteriza-se pelo início da condensação da cromatina. Isso se deve em grande parte à atuação de um complexo proteico denominado condensina. Um centrossomo inicia a formação do fuso mitótico Fragamentação do nucléolo: componentes em parte se dispersam pelo citoplasma na forma de ribonucleoproteínas e em parte permanecem associados a periferia dos cromossomos. Os dois centrossomos cada um com seu par de centríolos, começam a se mover para pólos opostos da célula e entre eles, pode se observar a formação de fibras polares (= microtúbulos) Cada um dos filamentos está constituído por duas cromátides (ditas irmãs). Coesina garante a coesão entre as cromátides-irmãs até o fim da metáfase.
 Cada um dos filamentos está constituído por duas cromátides (ditas irmãs). Coesina garante a coesão entre as cromátides-irmãs até o fim da metáfase.")
41
Cromatina mais condensada, mostrando filamentos mais grossos e mais curtos, e o nucléolo não é mais visualizado. Envoltório nuclear e as organelas membranosas, fragmentam-se em pequenas vesículas. 1. Contêm laminas B: permanecem associadas a membrana interna e laminas A: ficam livres no citosol. 2. Forma-se os cinetocoro, estrutura protéica ligada à região do centrômero de cada cromátide irmã, na qual os microtúbulos do fuso (cinetocóricos) se associam e exercem tensão sobre essas cromátides. 3. Remoção das das coesinas presentes entre os braços das cromátides, mas não das coesinas da região centromérica. 4. OBS: Nos fungos, as coesinas permanecem associadas ao longo de todo o comprimento do cromossomo até o final da metáfase.
 se associam e exercem tensão sobre essas cromátides. 3. Remoção das das coesinas presentes entre os braços das cromátides, mas não das coesinas da região centromérica. 4. OBS: Nos fungos, as coesinas permanecem associadas ao longo de todo o comprimento do cromossomo até o final da metáfase.")
42
As 3 classes de microtúbulos do fuso mitótico
astrais Microtúbulos cinetocóricos Microtúbulos interpolares

43
Cromatina atinge o máximo de condensação
1. Tensão proporcional que o s microtúbulos exercem em direção opostas sobre as cromátides-irmãs leva os cromossomos a assumir uma posição de equilíbrio em um plano na região equatorial da célula entre os dois pólos.

44
4. ANÁFASE Inicia-se repentinamente com a liberação da coesina que mantém as duas cromátides-irmã unidas. 3. Topoisomerase: enzima presente na região do cinetócoro, libera as fibras cromatínicas entrelaçadas para facilitar a separação das cromátides irmãs.

45
Começa abruptamente com a separação das cromátides-irmãs, que se movem para os pólos.
O posicionamento de cada homólogo do par independente um do outro no equador da célula permite que, ao separar as cromátides-irmãs, cada célula-filha receba todos os pares de cromossomos, mantendo assim a ploidia. O movimento das cromátides irmãs (cada uma agora denominada cromossomo-filho) para pólos opostos é resultante da combinação de dois processos: Anáfase A e Anáfase B.
 para pólos opostos é resultante da combinação de dois processos: Anáfase A e Anáfase B.")
46
. Nesse processo, ocorre o encurtamento dos microtúbulos do cinetocoro
. Os pólos das células se separam por alongamento dos microtúbulos polares

47
Se caracteriza pela reestruturação do envoltório nucleara a partir da reassociação dos componentes dispersos pelo citosol na pró-metáfase. . Os cromossomos irão se descompactar gradativamente até o final desta fase, assumindo o estado mais distendido da cromatina e característico da intérfase. . Os microtúbulos cinetocóricos desaparecem e os polares permanecem apenas na região equatorial, na qual se dará a citocinese.

48
Divisão citoplasmática da célula em duas, de maneira a assegurar que cada célula-filha receba um núcleo e quantidades suficientes dos constituintes celulares.

49
Animais anel contrátil (actina e miosina)
Citocinese Animais anel contrátil (actina e miosina)
")
50
Vegetais ocorre a formação de uma nova parede celular.

52
Mitose

53
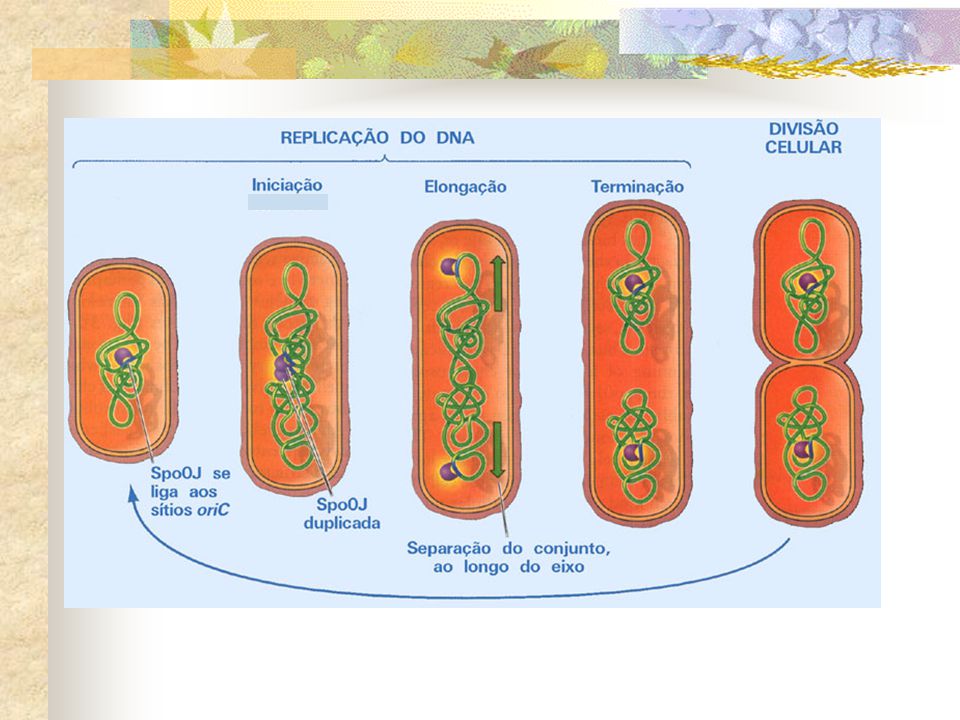
Divisão celular em procariotos
. Nos procariotos a divisão celular se dá por um processo denominado fissão binária, que é bastante simples comparado ao mecanismo observado nos eucariotos. . Antes da divisão física da célula, a molécula de DNA circular ancorada a membrana plasmática se replica. . Essa replicação ocorre de forma concomitante a um alongamento da célula e á uma expansão do volume da mesma.

55
As tarefas básicas do ciclo celular são realizadas de maneira bastante ordenada
O núcleo não se divide antes do término da síntese de DNA - E ao final de cada mitose, as células somáticas recém-formadas têm o mesmo tamanho de sua antecessora. Essas observações sugerem a existência de mecanismos de controle para as diferentes fases do ciclo.

56
3. Como elas coordenam toda a maquinaria que é necessária para esses dois processos?

57
O sistema de controle do ciclo celular funciona, de forma muito semelhante, ao sistema de uma máquina de lavar-roupas. Freios moleculares que podem parar o ciclo em vários pontos de checagem

58
Dois tipos de maquinaria estão envolvidos na divisão celular:
Uma produz os novos componentes da célula em crescimento; Atrai os componentes para os seus locais corretos e os reparte apropriadamente quando a célula se divide em duas. Qual o sistema de controle central que ativa e desativa toda essa maquinaria nos momentos corretos e coordena as atividades que produzirão o produto final?????

59
Na década de 1970, Y. Masui e D. Smith, trabalhando de forma independente, iniciaram um período intenso de descobertas científicas. Identificaram um fator que controlaria o início da divisão celular em oócitos anfíbios.

60
Oócitos em meiose contêm uma substância capaz de induzir a divisão celular em outros oócitos, ainda imaturos e extraídos cirurgicamente de fêmeas de sapos. Uma microinjeção com 5% do citoplasma em meiose era suficiente para que oócitos imaturos iniciassem o primeiro ciclo meiótico, semelhante ao que ocorre quando recebem diretamente o estímulo hormonal.

61
Fator promotor de maturação - MPF (maturation promoting factor)
Experimentos sugeriram: qualquer que fosse o fator causador da maturação, seria um componente do citoplasma das células em divisão por induzir a maturação dos oócitos, foi batizado de Fator promotor de maturação - MPF (maturation promoting factor) . Presente em todas as células em divisão estudadas, incluindo leveduras, células de invertebrados marinhos e de mamíferos. . Capaz de induzir mitose – fator promotor de mitose – mitosis promoting factor - MPF
 . Presente em todas as células em divisão estudadas, incluindo leveduras, células de invertebrados marinhos e de mamíferos. . Capaz de induzir mitose – fator promotor de mitose – mitosis promoting factor - MPF.")
62
Sempre antes da divisão celular
A atividade do MPF apresentava oscilações Durante intérfase O que poderia causar essa flutuação durante o ciclo celular?

63
Uma das pistas veio da seguinte constatação:
Divisões celulares podem ocorrer na ausência do próprio núcleo, mas não há progressão do ciclo sem síntese protéica; Imaginou-se que o MPF, ou algum de seus componentes, fosse sintetizado periodicamente durante o ciclo celular, promovendo a divisão da célula.

64
Intérfase Mitose Intérfase Mitose Intérfase Mitose Nível de Ciclina
Década de 1980, Tim Hunt et al. observaram alterações dramáticas nos níveis de síntese protéica em ovos de ouriço do mar após fertilização Nível de Ciclina Intérfase Mitose Intérfase Mitose Intérfase Mitose Tempo Ovos eram incubados com um aminoácido radioativo e, a cada 10 minutos, analisava-se o padrão das proteínas sintetizadas, marcadas radioativamente. - Maioria das proteínas se acumulava ao longo do ciclo, uma classe apresentava um padrão periódico, cíclico - ciclinas Acumulavam-se a cada intérfase, chegando ao máximo na transição metáfase-anáfase da mitose quando, em seguida eram abruptamente destruídas

65
Estrutura do MPF Ciclina B Dímero de ciclina B e proteína quinase Cdc2
– Lohka e Maller: purificaram alguns microgramas de MPF a partir de um litro de ovos de anfíbios: o MPF era formado por duas proteínas: Ciclinas e CdKs (quinases dependentes de ciclinas). Ciclina B Estrutura do MPF Dímero de ciclina B e proteína quinase Cdc2
. Ciclina B. Estrutura do MPF. Dímero de ciclina B e proteína quinase Cdc2.")
66
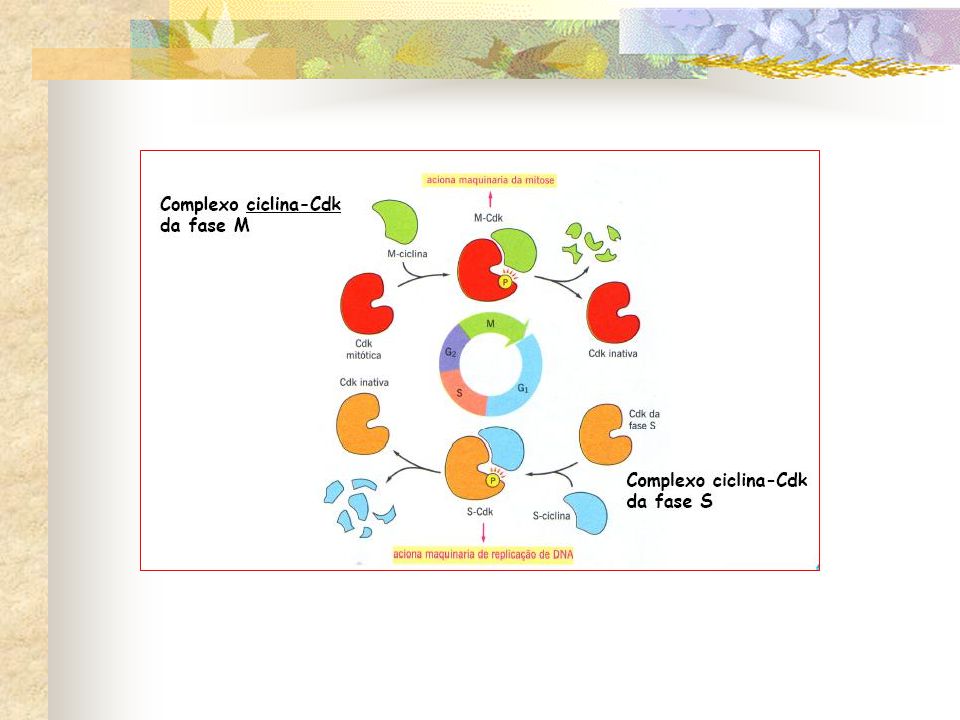
CICLINAS: proteínas transcritas e traduzidas em um determinado momento no ciclo celular (G1), especializadas na ativação de CdKs . CdKs: são proteínas que, ao se ligarem à ciclinas específicas, são ativadas, tornando-se capazes de adicionar grupos fosfato à certos resíduos de aminoácidos de proteínas alvo específicas, cuja fosforilação pode levar á uma alteração da atividade dessas proteínas alvo. CONTROLEADORES POSITIVOS DO CICLO CELULAR

68
Proteinoquinases dependentes de ciclinas são reguladas pelo acúmulo e pela destruição das ciclinas
Ciclinas são proteínas que passam por ciclos de síntese e degradação durante o ciclo celular. Dois tipos principais: Ciclinas mitóticas e ciclinas G1. CDK é constante ao longo do ciclo.

69
A diminuição súbita na concentração de M-ciclina no final da mitose é resultado da sua rápida destruição pelo sistema proteolítico dependente de ubiquitina Mitose se aproxima do término, múltiplas moléculas de proteína ubiquitina são ligadas as moléculas de ciclina-M

70
O que controla quando as ciclinas são ubiquitinadas e, dessa forma, marcadas para serem eliminadas?
Ciclina-M: Complexo Promotor de Anáfase (APC – anaphase promoting complex) adiociona ubiquitina à ciclina. > Não está ativa em todos os estágios do ciclo celular; > Atividade acionada no final da mitose
 adiociona ubiquitina à ciclina. > Não está ativa em todos os estágios do ciclo celular; > Atividade acionada no final da mitose.")
71
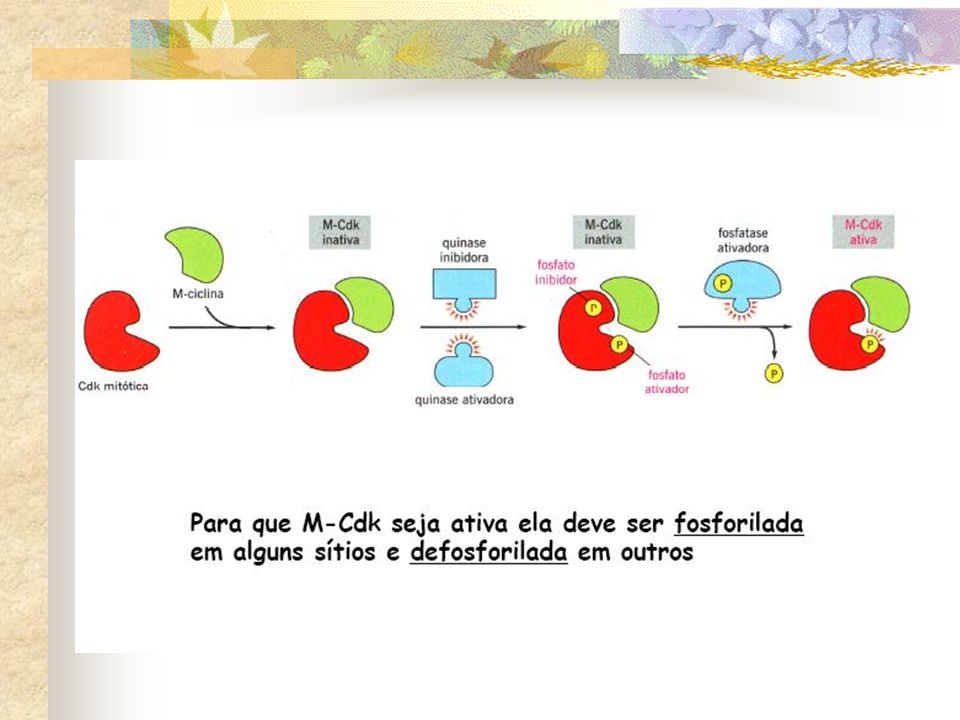
Atividade das Cdks também é regulada pela fosforilação e desfosforilação

72
O que aciona aciona essa rápida ativação de M-Cdk??????
No caso de M-Cdk, a concentração de M-ciclina aumenta gradualmente durante a intérfase, mas a atividade de M-Cdk é ativada subitamente no final da intérfase O que aciona aciona essa rápida ativação de M-Cdk??????

74
Essa retroalimentação positiva produz o súbito aumento explosivo na atividade de M-Cdk que conduz a célula repentinamente para a fase M.

75
Mecanismos de Regulação das Cdks
Associação com Ciclinas Ciclina Associação com inibidores de Cdk (CKIs) Fosforilação ativadora da treonina 160/161 Desfoforilação da treonina 14 e tirosina 15
 Fosforilação ativadora da treonina 160/161. Desfoforilação da treonina 14 e tirosina 15.")
78
Cdk inicia a replicação do DNA
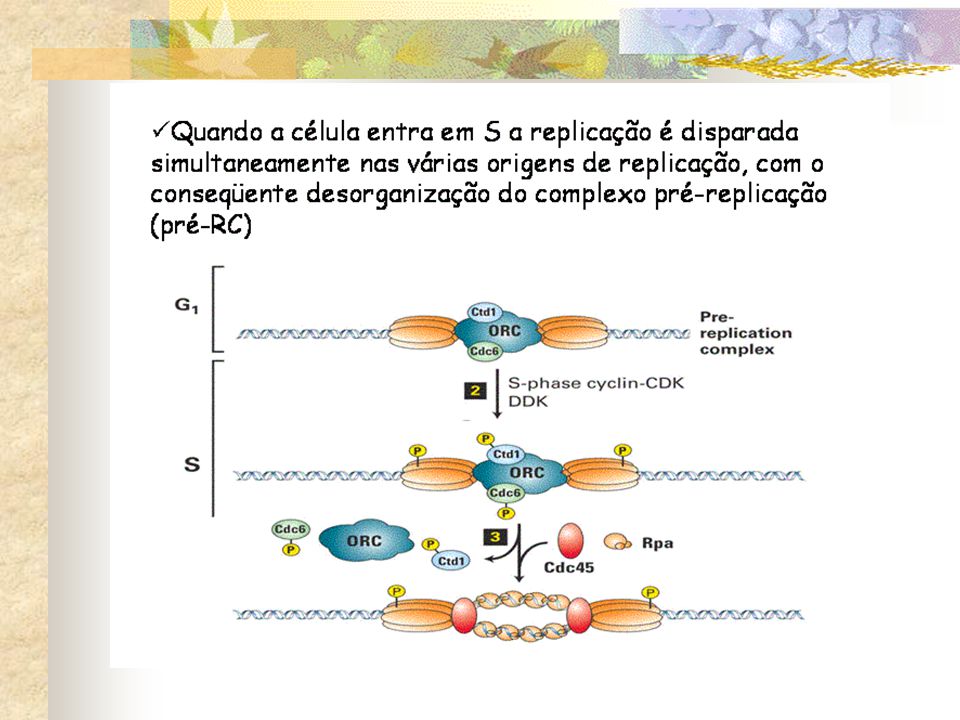
. A replicação do DNA inicia nas origens de replicação: sequencias nucleotídicas que estão dispersas em vários locais ao longo de cada cromossomo. Complexo multiprotéico – complexo de reconhecimento da origem (ORC: origin recognition complex) Permenece ligado as origens de replicação pelo ciclo celular, onde serve como um tipo de plataforma de aterrissagem para proteínas regulatórias adicionais que se ligam antes do início da fase S Quando se liga as ORCs em G1, ela promove a ligação de proteínas adicionais para formar o complexo pré-replicativo.
 Permenece ligado as origens de replicação pelo ciclo celular, onde serve como um tipo de plataforma de aterrissagem para proteínas regulatórias adicionais que se ligam antes do início da fase S. Quando se liga as ORCs em G1, ela promove a ligação de proteínas adicionais para formar o complexo pré-replicativo.")
80
Proteínas inibitórias do ciclo celular
Uma série de sinais fisiológicos é capaz de impedir a proliferação celular: a ligação entre a maquinaria do ciclo e os sinais que inibem a proliferação faz-se por proteínas inibitórias do ciclo celular: pontos de checagem. Proteínas capazes de inibir a atividade das CDKs 2 famílias p19 p18 Família INK4: interage exclusivamente com as CDK 4 e 6 p15 Família KIP (kinase inhibitor protein) inibe múltiplos complexos. Principal substrato para as cinases CDK4/6 é a pRB, esses inibidores teriam como função primordial modular indiretamente o estado de fosforilação dessa proteína supressora de tumor p57
 inibe múltiplos complexos. Principal substrato para as cinases CDK4/6 é a pRB, esses inibidores teriam como função primordial modular indiretamente o estado de fosforilação dessa proteína supressora de tumor. p57.")
81
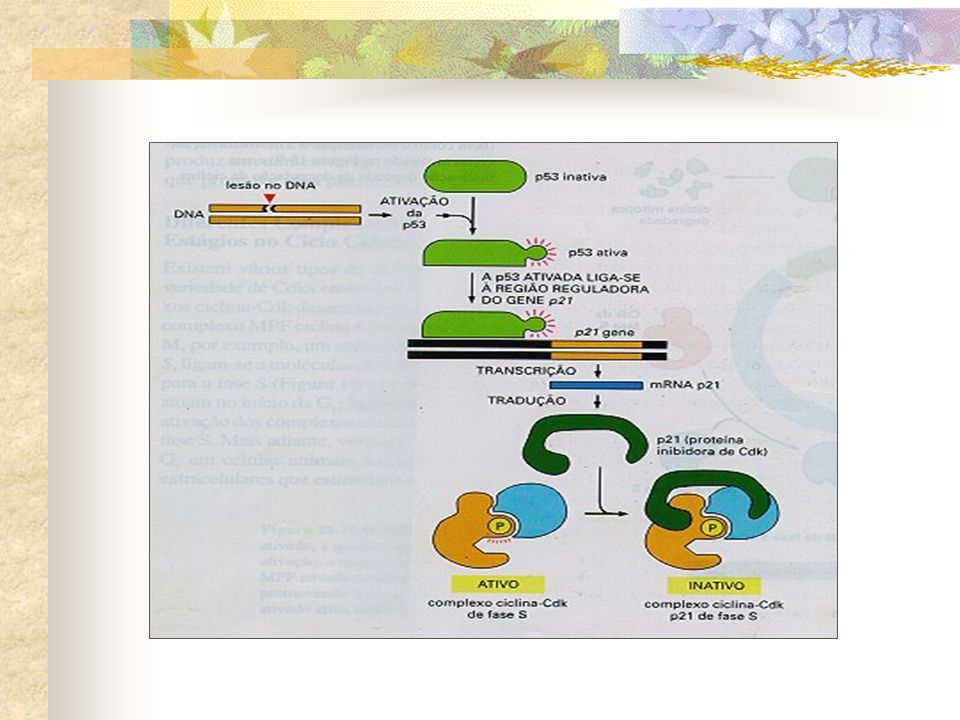
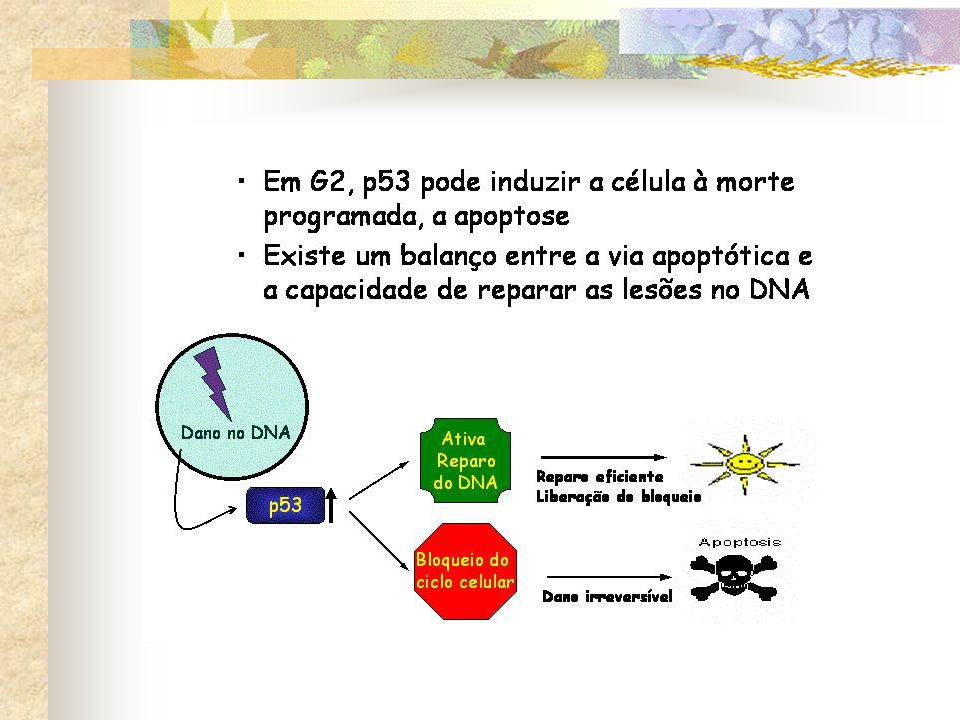
Radiação Dano no DNA Um dos mecanismos de ponto de checagem melhor compreendidos pára o ciclo celular em G1 se o DNA é danificado Aumento no nível de p53 Proteína reguladora de genes Parada em G1

84
Controle da transição metáfase-anáfase
. Ponto de checagem do fuso: garante um atraso na progressão do ciclo celular enquanto as condições para entrada na anáfase não forem alcançadas, assegurando a separação simultânea das cromátides-irmãs somente quando todos os cromossomos estiverem alinhados na placa metafásica com seus cinetocoros-irmãos ligados a microtúbulos de pólos opostos. Nesta fase, os cinetocoros que ainda não estabeleceram ligação com os microtúbulos recebem um complexo protéico contendo, dentre outras, as proteínas Mad2 e RubR1, que recrutam a proteína Cdc20 (ativadora do complexo promotor de anáfase – APC) para esse local, formando o chamado complexo de checagem. Estando aprisionada no complexo de checagem, Cdc20 não é capaz de ativar o APC, bloqueando, desse modo, o início da anáfase.
 para esse local, formando o chamado complexo de checagem. Estando aprisionada no complexo de checagem, Cdc20 não é capaz de ativar o APC, bloqueando, desse modo, o início da anáfase.")
85
. Quando ocorre a ligação de microtúbulos ao cinetocoro, o complexo de checagem se desfaz, liberando a Cdc20. . Essa proteína se liga ao APC ativando-o e possibilitando a degradação da securina, que por sua vez libera separase, que degradrá as coesinas, permitindo o desencadeamento da anáfase. A segregação não balanceada dos cromossomos pode levar à produção de células denominadas aneuplóides, que apresentam como conseqüência desse erro, cromossomos a mais ou a menos que o normal.

88
CÉLULAS ANIMAIS REQUEREM SINAIS EXTRACELULARES PARA SE DIVIDIR, CRESCER E SOBREVIVER
Para que uma célula animal se divida ou cresça, ou até mesmo sobreviva, apenas nutrientes não são suficientes – sinais químicos A maioria das moléculas de sinalização extracelulares são proteínas solúveis secretadas por outras células ou proteínas ligadas à superfície de outras células ou da matriz extracelular.

89
1. Mitógenos: estimulam a divisão celular, principalmente pela superação dos mecanismos de freio intracelulares que tendem a bloquear o avanço pelo ciclo celular 2. Fatores de crescimento: estimulam o crescimento celular 3. Fatores de sobrevivência: promovem a sobrevivência da célula pela supressão de apoptose.

90
Proteína Rb e o ponto de restrição
Retinoblastoma define um tipo de câncer infantil decorrente da inativação de ambos os alelos do gene Rb. . Cada célula tem duas cópias replicadas do gene do retinoblastoma (Rb). Quando as duas cópias do gene Rb sofrem mutação, uma proteína Rb anormal induz o crescimento cancerígeno de células da retina. . Quando uma única cópia do gene sofre mutação, a outra cópia do gene Rb funciona normalmente e suprime a proliferação celular irregular, a menos que ocorra uma segunda mutação. Proteína Rb e o ponto de restrição Proteína Rb como um inibidor do ciclo celular
. Quando as duas cópias do gene Rb sofrem mutação, uma proteína Rb anormal induz o crescimento cancerígeno de células da retina. . Quando uma única cópia do gene sofre mutação, a outra cópia do gene Rb funciona normalmente e suprime a proliferação celular irregular, a menos que ocorra uma segunda mutação. Proteína Rb e o ponto de restrição. Proteína Rb como um inibidor do ciclo celular.")
91
reprimida e não há síntese de DNA e progressão do ciclo celular.
Proteína Rb como um supressor O gene Rb codifica uma proteína nuclear envolvida na regulação da atividade de um grupo de proteínas – fatores de transcrição – envolvidos na síntese de DNA e na progressão do ciclo celular. Atividade gênica é reprimida e não há síntese de DNA e progressão do ciclo celular.

92
Proteína Rb fosforilada
Dissociação da Rb dos fatores de transcrição durante o final da fase G1 Fosforilação da Rb pelo complexo ciclina D-CdK4. Proteína Rb fosforilada Ligação dos fatores de transcrição aos genes-alvo

93
Aumentam a síntese e diminuem a degradação de macromoléculas

94
Em fim...... . Apesar dos detalhes do ciclo celular diferirem, dependendo do organismo e do estágio do desenvolvimento, os mecanismos centrais são os mesmos para todas as células. . Pode-se dizer que, de um modo geral, a síntese de ciclinas e a conseqüente ativação de CDK sempre resulta na fosfarilação de proteínas específicas que promovem a progressão do ciclo celular. . Alterações no microambiente celular e danos internos geram sinais intracelulares que influenciam o ciclo celular, justamente porque exercem controle sobre os complexos CDK acoplados a ciclinas.

Apresentações semelhantes












>")













