Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Ana Carolina Zakir Djanira Rodrigues Negrão João Paulo Apolari
UNIVERSIDADE DE SÃO PAULO CENA – CENTRO DE ENERGIA NUCLEAR NA AGRICULTURA Biodiversidade e abundância da microbiota do solo: ferramentas moleculares de estudo Ecologia Experimental de Micro-organismos Responsável: Pr0fª Drª Siu Mui Tsai Ana Carolina Zakir Djanira Rodrigues Negrão João Paulo Apolari

2
Introdução A estrutura e a atividade das comunidades microbianas do solo podem ser influenciadas por diversos fatores como: Composição e características físico-químicas; Formas de manejo e tipo de cultura empregada; Condições climáticas, geografia; Contaminações de natureza diversa: herbicidas, fungicidas, fertilizantes, metais pesados, poluentes orgânicos, etc;

3
Introdução

4
Independentes de cultivo
Introdução A microbiota do solo apresenta elevada diversidade genética e funcional, que pode ser caracterizada com base em estudos que empregam paralelamente: Dependentes de cultivo Independentes de cultivo

5
Introdução Técnicas independentes de cultivo: T-RFLP;
Construção de biblioteca do gene 16S rRNA; Clonagem e sequenciamento de genes, como o nifH; Análise filogenética; q-PCR. RT-RFLP.

6
Estudo de caso 1

7
Objetivos Análise da biodiversidade microbiana do solo da rizosfera
de cana-de-açúcar com adubação de fertilizante nitrogenado.

8
Amostragem Cana-de-açúcar: RB72454 Tratamentos: com e sem adubação.
Controle: mesmo local, ano anterior com adubação. Dois diferentes regimes de fertilização: Sem fertilizante nitrogenado. 120 kg/ha de fertilizante nitrogenado. O controle utilizado foi uma amostra do solo de rizosfera de cana-de-açúcar crescida no mesmo local no ano anterior, que também recebeu adubação nitrogenada.

9
Amostragem O solo aderido às raízes foi coletado por agitação manual em tubos e imediatamente armazenado a -70 °C para análise. Um total de 1 g de solo de cada amostra foi utilizado para a extração de ácido nucleico.

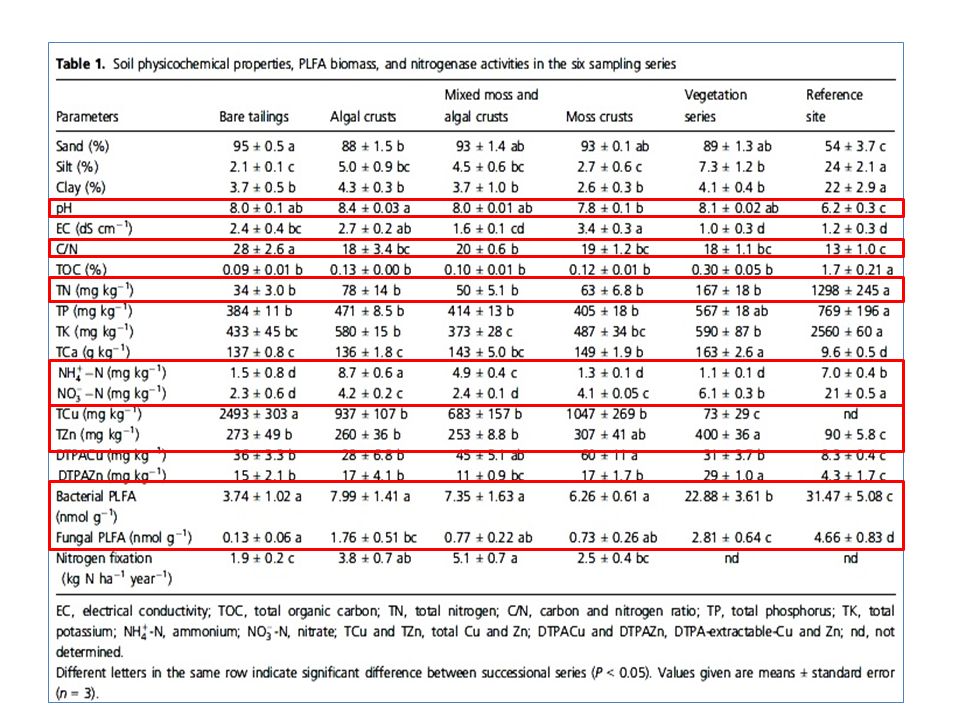
10
Extração do DNA O kit UltraClean soil DNA isolation
(Mobio Laboratories, inc., USA).
.")
11
AMPLIFICAÇÃO 16S O gene 16S rRNA foi amplificado por PCR.
Os primers: 27F (5’AGAGTTTGATCMTGGCTCAG3’) e 1492r (5’GGYTACCTTGTTACGACTT3’) foram usados para amplificar fragmentos de aproximadamente 1500 pb.
 e 1492r (5’GGYTACCTTGTTACGACTT3’) foram usados para amplificar fragmentos de aproximadamente 1500 pb.")
12
Construção de biblioteca de 16S rRNA e sequenciamento
Amplificação 16S rRNA Clonagem PCR2.1-TOPO (Invitrogen, USA) Seleção de 12 clones por placa (96 poços) Digestão EcoRI Sequenciamento dos fragmentos
 Seleção de 12 clones por placa (96 poços) Digestão EcoRI. Sequenciamento dos fragmentos.")
13
Sequências e análise de diversidade
Remoção das sequências de baixa qualidade (Phred base caller) Taxonomia (Ribossomal Database Project) Alinhamento de sequências (Mothur v.1.9.0) Depósito de Sequência no banco de dados (≥ 50 pb) (GenBank database) MEGA-BLAST (Busca de Sequências depositadas no NCBI)
 Taxonomia (Ribossomal. Database Project) Alinhamento de sequências. (Mothur v.1.9.0) Depósito de Sequência no banco de dados (≥ 50 pb) (GenBank database) MEGA-BLAST. (Busca de Sequências. depositadas no NCBI)")
14
RESULTADOS

15
Construção de biblioteca de 16S rRNA e sequenciamento
S0N*= solo de rizosfera de plantas com 12 meses (1 ano antes) S13N*= solo de rizosfera de plantas com 13 meses (1 ano depois) S13n= solo de rizosfera de plantas com 13 meses (1 ano depois) Foram obtidas um total de 541 sequências com mais de 50 pb 390 (72,1 %) dessas sequências tiveram mais de 250 pb
 S13N*= solo de rizosfera de plantas com 13 meses (1 ano depois) S13n= solo de rizosfera de plantas com 13 meses (1 ano depois) Foram obtidas um total de 541 sequências com mais de 50 pb. 390 (72,1 %) dessas sequências tiveram mais de 250 pb.")
16
Distribuição taxonômica dos clones das biblioteca de 16S rRNA
86 % das sequências poderiam ser atribuídas em 15 filos. 127 (23,6 %) foram atribuídas a nível de gênero com confiança média de 95,5% nas 3 bibliotecas. vc vc vc vc
 foram atribuídas a nível de gênero com confiança média de 95,5% nas 3 bibliotecas. vc. vc. vc. vc.")
17
Para investigar a classificação destes clones, as sequências foram recuperadas e submetidas a uma pesquisa MEGA-BLAST na base de dados NCBI. Identidades elevadas (98 a 100%) com bactérias não cultivadas e / ou Bacillus isolados a partir de solo agrícola contaminado.
 com bactérias não cultivadas e / ou Bacillus isolados a partir de solo agrícola contaminado..")
18
Riqueza de comunidade pelo índice de Chao:
1º - S13N (298), 2º - S0N (220), 3º - S13n (217) Diversidade das bibliotecas pelo índice de Shannon: 1º - S13N (4,25), 2º - S0N (4,19), 3º - S13n (4,09). No entanto, as três amostras apresentaram riqueza e diversidade semelhante, sem diferença significativa no intervalo de confiança de 95%.
, 2º - S0N (220), 3º - S13n (217) Diversidade das bibliotecas pelo índice de Shannon: 1º - S13N (4,25), 2º - S0N (4,19), 3º - S13n (4,09). No entanto, as três amostras apresentaram riqueza e diversidade semelhante, sem diferença significativa no intervalo de confiança de 95%.")
19
CONCLUSÃO Esperava-se que nas amostras S0N e S13N que receberam adubação nitrogenada as populações bacterianas compartilhariam grupos comuns. O que de fato não aconteceu, sugerindo que a adubação nitrogenada pode não ser o principal fator que influencia a biodiversidade do solo.

20
CONCLUSÃO O número de sequências de Verrucomicrobia recuperados à partir de ambas amostras com fertilizantes foram significativamente menores em relação às amostras sem fertilizantes (S13n), sugerindo que este grupo pode ser um indicador de solos ricos em nitrogênio.
, sugerindo que este grupo pode ser um indicador de solos ricos em nitrogênio.")
21
Estudo de caso 2

22
OBJETIVOS Estudar a diversidade e abundância de bactérias fixadoras de N2, através do gene nifH, em resíduos de mina de cobre. Explorar o papel determinante na distribuição das comunidades diazotróficas na sucessão primária.

23
Local dos resíduos de cobre em Yangshanchong

24
Características da área de estudo
Mais importante fonte de cobre na China. Precipitação anual: 1300 mm. Temperatura média anual: 16 ºC. Operações durante 30 anos, abandonada em 1990. Área aproximada de 20 ha. Monitoramento da área durante sete anos.

25
AMOSTRAGEM Resíduos sem cobertura. Agregados biológicos de algas.
Agregados biológicos de algas e musgo. Agregados biológicos de musgo. Comunidade de plantas vasculares. Sítio de referência – local distante dos resíduos.

26
AMOSTRAGEM Coleta: 0 – 10 cm, com sonda (3 cm x 20 cm).
Três replicatas (compostas de 5 sondas cada). Mantidas sob refrigeração. Homogeneizadas e peneiradas (2 mm). Para as análises moleculares: - 40ºC.
. Mantidas sob refrigeração. Homogeneizadas e peneiradas (2 mm). Para as análises moleculares: - 40ºC.")
27
ANÁLISES pH. Condutividade elétrica. NO3- e NO4-
Carbono orgânico total (TOC-VCPH). Nitrogênio total (Kjeltec TM 2300). Concentrações de Cu e Zn (Page et al., 1982). Metais pesados (IVP-OES; OPTIMA 2100).
. Nitrogênio total (Kjeltec TM 2300). Concentrações de Cu e Zn (Page et al., 1982). Metais pesados (IVP-OES; OPTIMA 2100).")
28
Análise da biomassa microbiana Atividade da nitrogenase
ANÁLISES Análise da biomassa microbiana Lipídios totais: 5 g de solo. Frações polares de ácidos graxos de fosfolipídios: separadas, quantificadas, identificadas por cromatografia gasosa. Atividade da nitrogenase Realizada in situ. Valores médios obtidos foram convertidos em input de fixação biológica de N2.

29
Extração do DNA Análises de T-RFLP
DNA total da comunidade microbiana: 0,5 g de solo/amostra. Amplificação da região do gene nifH (365 pb) com os primers: PolF e PolR (Poly et al., 2001). Primer PolF foi rotulado em 5’ com FAM (6-carboxifluoresceina).
 com os primers: PolF e PolR (Poly et al., 2001). Primer PolF foi rotulado em 5’ com FAM (6-carboxifluoresceina).")
30
Triplicata de PCR por amostra Extração do DNA Análises de T-RFLP
Purificação Digestão: 37 ºC/6 h Enzima HaeIII. Eletroforese ABI 3730xl Os tamanhos dos fragmentos e a intensidade de fluorescência foram analisados pelo software GENESCAN. Perfil de RFLP

31
Análises de T-RFLP A abundância relativa dos fragmentos de restrição terminal individual (T-RFs) foi calculada pela % do perfil da altura dos picos. Apenas os T-RFs com abundância relativa > 1% foram analisados.

32
Clonagem do gene nifH As bibliotecas de clones foram construídas das amostras: Resíduos SC, alga+musgo, musgo, sítio de referência. Os fragmentos de nifH foram amplificados (igual à extração), com primers sem fluoróforos. Purificação das triplicatas de PCR. Clonagem de nifH, com o kit TOPO TA.
, com primers sem fluoróforos. Purificação das triplicatas de PCR. Clonagem de nifH, com o kit TOPO TA.")
33
Clonagem do gene nifH Amplificação das inserções dos clones de nifH recombinantes Digestão enzimática dos clones com endonucleases HhaI e HaeIII Determinação em eletroforese (gel de agarose, 2%) Comparação manual do resultado do perfil de RFLP Os clones representantes de cada padrão único de RFLP foram sequenciados (ABI 3730xl) As UTOs foram definidas com pelo menos 95 % de sequências de nucleotídeos similares (sotware DOTUR)
 Comparação manual do resultado. do perfil de RFLP. Os clones representantes de cada padrão único de RFLP foram sequenciados (ABI 3730xl) As UTOs foram definidas com pelo menos 95 % de sequências de nucleotídeos similares (sotware DOTUR)")
34
Análise filogenética Comparação das sequências de nifH no GenBank, BLAST. Tradução das sequências em aminoácidos (MEGA 3.1). Construção da árvore filogenética, usando distâncias de correção Poisson.

35
Análises de q-PCR A abundância do gene nifH foi quantificada.
Primers idênticos ao RFLP. Reações padrão (LightCycler 480). Confirmação em gel de agarose. Controles negativos sem DNA. Análises estatísticas: correlação entre densidade de nifH e propriedades físico-químicas do solo.
. Confirmação em gel de agarose. Controles negativos sem DNA. Análises estatísticas: correlação entre densidade de nifH e propriedades físico-químicas do solo.")
36
Depósito das sequências de nucleotídeos
As sequências nifH foram depositadas: EMBL/GenBank/DDBJ Número de acesso: FN FN

37
RESULTADOS

39
Índices de Diversidade das Comunidades Diazotróficas
Total de T-RFs em todos os perfis: 45 (3 comuns, 9 únicas), média = 13. Diversidade de nifH indicado pelos três índices calculados: não mostrou grande significância (+/-) nos locais com resíduos.
, média = 13. Diversidade de nifH indicado pelos três índices calculados: não mostrou grande significância (+/-) nos locais com resíduos.")
40
Riqueza e Diversidade das sequências de nifH

41
* Análises comparativas de sequências de 204 clones nifH das 4 bibliotecas revelaram filotipos únicos. * A maioria distribuída em Proteobacteria, Cyanobacteria e Firmicutes. * 44% das UTOs foram relacionados à bactérias desconhecidas, representando novos filotipos.

42
Sequências de nifH filiadas a Cyanobacteria foram detectadas somente nos solos com ABio, constituindo 23-39% das bibliotecas de clones.

43
A abundância relativa de nifH pode ser correlacionada com o progresso da sucessão microbiana e da atividade da nitrogenase. 5.06 x 105

44
Resultados Houve correlação positiva entre TOC, TN, NO3- com o número de cópias de nifH. A relação C/N teve correlação negativa com a abundância de nifH. O pH e os metais pesados mostraram efeitos negativos sobre a abundância de nifH.

45
resíduos nus agregados biológicos
CONCLUSÕES Análises de T-RFLP revelaram diversidade crescente: resíduos nus agregados biológicos Nenhuma UTO encontrada foi comum para todas as amostras, indicando que o progresso da sucessão pode abrir novos nichos. A sucessão da colonização nos resíduos de minas de cobre geralmente é acompanhada por mudanças nas suas características físico-químicas.

46
CONCLUSÕES A natureza heterogênea dos resíduos pode ser afetada pela idade e presença de plantas pioneiras. Variações nas condições ambientais podem influenciar a diversidade e abundância de microrganismos indígenas, inclusive os envolvidos no Ciclo do nitrogênio. Esses fatores podem afetar positivamente a sucessão primária de comunidades diazotróficas em locais altamente perturbados.

47
Estudo de caso 3

48
Objetivos Identificar relações importantes entre a comunidade bacteriana do solo e seu meio ambiente. Comparar a dinâmica da estrutura da comunidade bacteriana e da atividade ao longo três épocas diferentes em dois locais geograficamente distintos, alpes sem vegetação e local de geleira. Destacar mudanças nos parâmetros climáticos, concentrações nutricionais, e as respostas da comunidade bacterianas (estrutura e atividade), que podem ocorrer durante sucessão sazonal.
, que podem ocorrer durante sucessão sazonal.")
49
Local da coleta Damma Tsanfleuron Solo coletado em duas áreas geologicamente, química e biologicamente diferentes: duas geleiras alpinas, Suíça.

50
AMOSTRAGEM 10 subamostras Coletas em de 0 - 5cm maio – out/2008
Três estações: primavera (I), verão (II) e outono (III) Peneiradas (2 mm) coletados em sacos plásticos Inverno não coletou Alíquotas de 2 ml (tubos eppendorf) em gelo seco CLIMAP Alíquotas foram armazenadas a -20°C para extração de DNA, e a -80°C para extração de RNA.
, verão (II) e outono (III) Peneiradas (2 mm) coletados em sacos plásticos. Inverno. não coletou. Alíquotas de 2 ml (tubos eppendorf) em gelo seco. CLIMAP. Alíquotas foram armazenadas a -20°C para extração de DNA, e a -80°C para extração de RNA.")
51
Análises do Solo Umidade: secagem em estufa por 3 dias, 60 °C.
Nutrientes: cromatógrafo de íons (IC DIONEX 2300). NH4+ e PO43-: colorimetricamente (Mulvaney, 1996). pH: através de CaCl2 (Mettler Toledo XMP225). DOC: analisado por Shimadzu SSI TOC-5000. TC e TN: determinados por combustão (Leco 932CHNS).
. NH4+ e PO43-: colorimetricamente (Mulvaney, 1996). pH: através de CaCl2 (Mettler Toledo XMP225). DOC: analisado por Shimadzu SSI TOC TC e TN: determinados por combustão (Leco 932CHNS).")
52
Análises Enzimáticas FDA (Fluorescein diacetate assay, Rosswall, 1982). Atividades potenciais de enzimas por meio do ensaio de MUB (Classen et al., 2006) com modificações: Fosfatase alcalina. Quitinase (N-acetil-glucosaminidase). Sulfatase. β-glucosidase.
 com modificações: Fosfatase alcalina. Quitinase (N-acetil-glucosaminidase). Sulfatase. β-glucosidase.")
53
Biomassa e Extração do DNA
Número de células: Coloração DAPI (Bennett et al., 2006). Concentração de DNA: utilizada como substituto para biomassa microbiana (Marstorp e Witter, 1999). Extração do DNA: por kit Zymo microbial DNA. Quantificação do DNA: de acordo com Sandaa et al. (1998), através de curvas de valores de fluorescência (0 a 100 ng µl-1).
. Concentração de DNA: utilizada como substituto para biomassa microbiana (Marstorp e Witter, 1999). Extração do DNA: por kit Zymo microbial DNA. Quantificação do DNA: de acordo com Sandaa et al. (1998), através de curvas de valores de fluorescência (0 a 100 ng µl-1).")
54
Análises Moleculares T-RFLP Extração do DNA PCR com primers marcados
Sequenciamento RT-T-RFLP Extração RNA Síntese de cDNA Transcrição reversa: SSII transcriptase reversa, com iniciador UNI-b-rev (Bundt et al., 2001). PCR Sequenciamento
. PCR. Sequenciamento.")
55
Biblioteca de clones de cDNA e sequenciamento
Três bibliotecas: um ponto por estação Purificação de cDNA Qiagen PCR Purification Reação pGEM T-Easy vector Transformação em células JM109 (Promega) Eletroforese Purificação das reações positivas ( Qiagen 96) Sequenciamento Definição dos filotipos
 Eletroforese. Purificação das reações positivas ( Qiagen 96) Sequenciamento. Definição dos filotipos.")
56
Depósito das sequências
GenBank Número de acesso: HM HM623763

57
Resultados

58
Condições Climáticas

59
Propriedades de Solo

60
Atividade Enzimática Específica

61
Biomassa Microbiana

62
Biomassa Microbiana

63
Estrutura da Comunidade Microbiana

64
Estrutura da Comunidade Microbiana

65
Estrutura da Comunidade Bacteriana Viável

66
Estrutura da Comunidade Bacteriana Viável
Grupo mais abundante: α-Proteobacteria. Gêneros Methylobacterium sp. (AB252210) e Methylobacterium aquaticum (AB252197), com sequências > 97 %. Actinobacteria: 39,2 % - outono de Damma. Actinobacteria: 39,1 % - verão de Tsanfleuron. Corynebacterium tuscaniensis (AY677186) foi o mais comum filotipo de Actinobacterium encontrado. Firmicutes: frequente em ambos os locais (> 28 % do verão Damma, do outono Tsanfleuron).
 e Methylobacterium aquaticum (AB252197), com sequências > 97 %. Actinobacteria: 39,2 % - outono de Damma. Actinobacteria: 39,1 % - verão de Tsanfleuron. Corynebacterium tuscaniensis (AY677186) foi o mais comum filotipo de Actinobacterium encontrado. Firmicutes: frequente em ambos os locais (> 28 % do verão Damma, do outono Tsanfleuron).")
67
Estrutura da Comunidade Bacteriana Viável
Gênero Paenibacillus (amostras Damma). Gêneros Bacillus e Paenibacillus (amostras Tsanfleuron). Interessantemente, as bibliotecas de clones da primavera apresentou uma variedade maior de filotipos em comparação com estações seguintes: Primavera Damma biblioteca, Rhodospirillales (e.g. DQ094180) estavam entre os mais abundantes filotipos de α-Proteobacteria detectados.
. Gêneros Bacillus e Paenibacillus (amostras Tsanfleuron). Interessantemente, as bibliotecas de clones da primavera apresentou uma variedade maior de filotipos em comparação com estações seguintes: Primavera Damma biblioteca, Rhodospirillales (e.g. DQ094180) estavam entre os mais abundantes filotipos de α-Proteobacteria detectados.")
68
Resultados Poucos estudos são realizados considerando-se as relações entre variações físico-químicas do solo e das comunidades bacterianas em ambientes áridos/estéreis. A estrutura das comunidades bacterianas no verão apareceram relacionadas principalmente às variáveis climáticas.

69
Resultados Comunidades de amostras nas estações da primavera e do outono foram relacionadas à variações na composição de nutrientes. α-Proteobacteria foi a linhagem dominante ao longo de todo o período de amostragem, sugerindo que populações bacterianas viáveis em ambientes extremos são geralmente capazes de sustentar fortes oscilações ambientais.

70
Obrigado!!!

Apresentações semelhantes
>")



 dos genes e seu armazenamento>")





>")








