Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Daniel Nizzo Emílio Telles Guilherme Mattos Guilherme Ribeiro
Larissa Guimarães Raul Leal Virologia 2011/2 Prof Mônica Freitas

2
INTRODUÇÃO ● O vírus Influenza A é um patógeno originado em animais não humanos mas que pode ser transmitido à eles, além de circular e mudar constantemente suas características; ● Infecção com 2 tipos de Influenza A pode levar a mistura de segmentos do genoma e resultar em um novo tipo de vírus (principal causa de epidemia); ● A evolução do vírus Influenza ocorre por recombinação gênica, gerando um novo subtipo viral, e por mutação causada pela seleção do vírus mediada por anticorpos.
; ● A evolução do vírus Influenza ocorre por recombinação gênica, gerando um novo subtipo viral, e por mutação causada pela seleção do vírus mediada por anticorpos.")
3
INTRODUÇÃO Classificação do vírus Influenza A em dois grandes grupos de acordo com as propriedades antigênicas do HA

4
INTRODUÇÃO O genoma do vírus Influenza A consiste em 8 ssRNA (-)
que traduzem em 11 proteínas

5
Ciclo de replicação do vírus Influenza A
INTRODUÇÃO Ciclo de replicação do vírus Influenza A

6
A GRIPE NO SÉCULO 21 Epidemia – “é doença geralmente infecciosa, de caráter transitório, que ataca grande número de indivíduos em uma determinada localidade.” Pandemia – “é enfermidade epidêmica amplamente disseminada.” Endemia – “é doença infecciosa que ocorre habitualmente e com incidência significativa em dada população ou região.” O potencial de pandemia do vírus H5N1, assim como os numerosos surtos em aves selvagens, ainda tem criado uma atenção sobre o vírus da gripe aviária em várias regiões ao redor do mundo. No entanto, o surgimento da pandemia de H1N1 em 2009, da gripe suína, revelou a falta de vigilância sistemática de outros hospedeiros suscetíveis. Esforços em grande escala na vigilância desses casos têm sido ajudados pelo desenvolvimento de várias tecnologias-chaves, usadas para obter seqüenciamento completo do genoma viral, selvagens e clínicos. Uso de uma grande bases de dados de seqüências filogenética e ferramentas de análise, estão permitindo estudos epidemiológicos mais rápidos e abrangentes sobre o vírus influenza em reservatórios humanos e naturais.

7
Vigilância dos vírus aviários
Vírus da H5N1 surgiu a partir de mutações no sítio de clivagem HA provavelmente em aves selvagens afetando também as doméstica. Vários surtos de vírus da H5N1 ocorreram em aves domésticas e o primeiro caso em humano foi documentado em 1997. No geral foram 562 casos humanos de 15 países, com uma taxa de mortalidade de ~ 59% (329 mortes). E até agora a maioria dos casos têm sido associados com direto contato humano com espécies de aves infectadas. A Vigilância das aves selvagens é a chave para a compreensão da origem patogênica, evolução e disseminação mundial desses vírus. Desde 2002, o genótipo Z (pequenas deleções nos genes que codificam as proteínas NS1 e NA) tem sido o genótipo predominante no H5N1. No entanto, dez principais linhagens distintas da H5N1 vírus foram identificados até a data, demonstrando a evolução destes vírus na natureza. Abrangentes estudos genômicos de um conjunto diversificado dos vírus da gripe aviária (AIVs), demonstraram que a maior variabilidade encontra-se nas proteínas, HA , NA e NS1 , e levou à descoberta de um suposto PDZ ligante no domínio terminal da NS1 que estar envolvidos em virulência.
. E até agora a maioria dos casos têm sido associados com direto contato humano com espécies de aves infectadas. A Vigilância das aves selvagens é a chave para a compreensão da origem patogênica, evolução e disseminação mundial desses vírus. Desde 2002, o genótipo Z (pequenas deleções nos genes que codificam as proteínas NS1 e NA) tem sido o genótipo predominante no H5N1. No entanto, dez principais linhagens distintas da H5N1 vírus foram identificados até a data, demonstrando a evolução destes vírus na natureza. Abrangentes estudos genômicos de um conjunto diversificado dos vírus da gripe aviária (AIVs), demonstraram que a maior variabilidade encontra-se nas proteínas, HA , NA e NS1 , e levou à descoberta de um suposto PDZ ligante no domínio terminal da NS1 que estar envolvidos em virulência.")
8
Vigilância dos vírus aviários
A Vigilância das Aves selvagens da América do Norte indicou que suas cepas e da Eurásia raramente se misturam e não detectaram evidência de vírus H5N1. No entanto, vírus isolados do Estados Unidos demonstraram uma alta taxa de recombinação. Isso pode ser um sinal de uma mistura constante de genomas virais em vez de disseminação de um conjunto constante de segmentos que caracteriza a influenza A em mamíferos. Em 2005, um vírus da H5N1 foi responsável por um surto em aves aquáticas no lago Qinghai, oeste da China,resultando em alta mortalidade de aves e levantando preocupações do potencial de transmissão entre aves migratórias. Posteriormente, geneticamente e antigenicamente, cepas distintas H5N1 apareceram ao longo do território Sudeste da Ásia e tornaram-se endêmicas. Mais recentemente, linhagens virais H5N1 surgiram através de rearranjo com as endêmicas que estão presentes em aves aquáticas locais, levando a seleção de vírus que são capazes de infectar múltiplas espécies aviária, permitindo a disseminação geográfica.

9
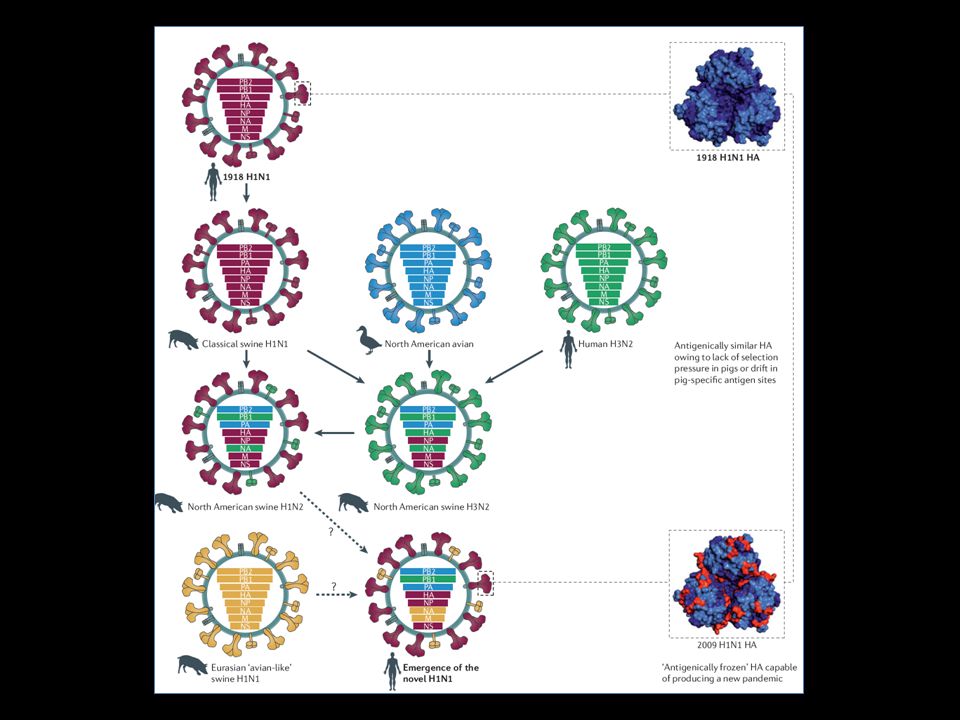
Gripe Suína H1N1 A gripe 2009 H1N1 possivelmente surgiu em abril de 2009 na América do Norte (México). Seu genoma sugeria que não havia marcadores previamente conhecido de adaptação humana, e por isso seus antecessores não detectados, circularam em suínos por algum tempo até o rearranjo e da detecção do vírus em seres humanos. Primeiros dados epidemiológicos indicaram que a estimativa de reprodução do vírus seria semelhante aos de pandemias de gripe anteriores, e que em crianças <15 anos idade a taxa infecção foi o dobro que em adultos. Pouco depois de surtos em todo o mundo, começou uma dinâmica propagação do vírus que foi rapidamente rastreadas e mapeadas. Surtos foram caracterizados por várias Regiões indicando uma pandemia. A dispersão geográfica ocorreu em três etapas principais: primeiro - propagação do México para os Estados Unidos; de transmissão segundo - sustentado na América do Norte, uma disseminação inicial a outras partes do mundo terceiro - distribuídos globalmente e focos secundários fora das Américas. Estudos demonstraram que o vírus circulou na população humana por um período de até 3 meses antes de ser identificado, isto é, a data da primeira infecção. (estimado entre 29 de dezembro de 2008 e 22 de Fevereiro 2009) Em estudo realizado no sul da China, mostrou que a recombinação entre as extensas linhagens de influenza suína e humano-aviárias (como mostrada FIG. 1b e 2) levou à surgimento de diferentes vírus com antígeno e genes de gripe suína em cerca de 2007. O potencial para mais rearranjo do vírus da pandemia H1N1 2009, força uma vigilância mais sistemática e abrangente de todo o mundo nos porcos para caracterizar e detectar cepas circulantes virais com um risco potencial para os humanos.
 Em estudo realizado no sul da China, mostrou que a recombinação entre as extensas linhagens de influenza suína e humano-aviárias (como mostrada FIG. 1b e 2) levou à surgimento de diferentes vírus com antígeno e genes de gripe suína em cerca de O potencial para mais rearranjo do vírus da pandemia H1N1 2009, força uma vigilância mais sistemática e abrangente de todo o mundo nos porcos para caracterizar e detectar cepas circulantes virais com um risco potencial para os humanos.")
10
Epidemiologia molecular sazonal do vírus humano
Os vírus sazonais H1N1 e H3N2 vem circulando desde 1977. Algumas análises sobre o sequenciamento dos vírus H1N1 e H3N2, permitiram a postulação de um modelo para explicar a origem da tensão sazonal anual, propondo que “nos trópicos, a população sofre uma forte seleção antigênica e serve como fonte para epidemia do vírus influenza, semelhante à vírus exportados diretamente da sua origem populacional nos hemisférios norte e sul.

11
Este “review” apresenta essa sobreposição epidêmica no oeste e sudeste asiático gerando uma circulação contínua do vírus influenza H3N2 dentro dessa região, e se espalhando para a Oceania, América do Norte, Europa e subsequentemente para a América do Sul. A partir daí, as semelhanças encontradas entre os vírus do oeste e sudeste asiático podem ajudar a determinar as características antigênicas do vírus que, posteriormente, devem circular pelo resto do mundo. Em acordo com essas informações , a emergência do vírus influenza, causador da epidemia sazonal, é largamente influenciada pela migração da população mundial e não resultado de vírus sendo reativados em hospedeiros durante o inverno.

12
As condições do inverno aparecem para contribuir na eficiência da transmissão e extensão do vírus influenza. De maneira adversa, a emergência pandêmica do vírus H1N1 em 2009 originou-se no hemisfério norte durante a primavera e se espalhou durante o verão, sendo a segunda onda de infecção em massa antes do outono, indicando que embora as condições climáticas influenciem a epidemiologia do vírus influenza A, sua transmissão e extensão pode ocorrer de maneira eficiente em populações “novas”.

13
Emergência pandêmica do vírus influenza H1N1 em 2009
O vírus influenza H1N1 possivelmente emergiu em Abril, através da introdução em humanos na América do Norte (mais precisamente no México). Seu genoma não possui nenhuma das adaptações previamente conhecidas, dando ao vírus um aspecto de “novo”. Seu antepassado, possivelmente, foi a circulação não-detectada em porcos durante um certo período de tempo sugerindo a proximidade com os precursores suínos detectados no genoma do influenza.
. Seu genoma não possui nenhuma das adaptações previamente conhecidas, dando ao vírus um aspecto de novo . Seu antepassado, possivelmente, foi a circulação não-detectada em porcos durante um certo período de tempo sugerindo a proximidade com os precursores suínos detectados no genoma do influenza.")
14
Análises evolucionária e genética revelaram que esse vírus contém um complexo genoma, originado de vírus que infectam pássaros, humanos e suínos, e que possivelmente derivam de vírus que circularam em populações suínas e não foram identificadas por aproximadamente uma década.

16
Novos conceitos em tropismo hospedeiro
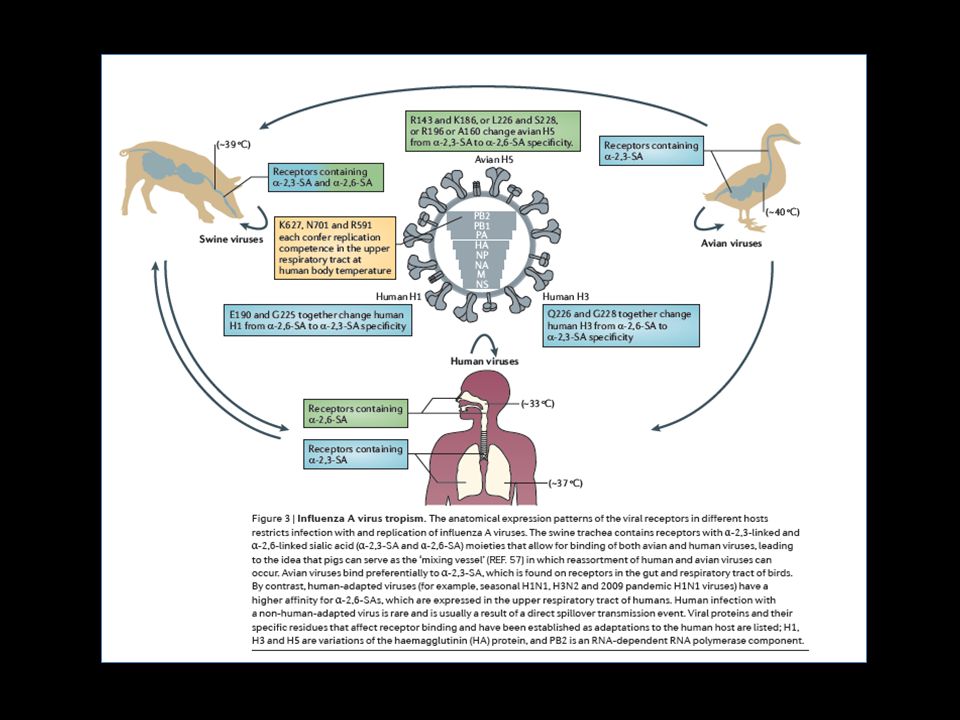
Relação vírus x hospedeiro - especificidade - infecção esporádicas e barreiras do hospedeiro Taxia tecidual e especificidade de receptor A especificidade e afinidade da hemaglutinina viral pelo seu receptor é crucial. ex. mutação do H1N1 associada a dupla especificidade e morte e H1N1 sazonal com adaptação ao hospedeiro, adaptações específicas necessárias; Humanos = receptores α - 2,6 - SA (sazonal) e α - 2,3 - SA (aviário), ex. H1 e H3; Aves = galactose presente nos receptores α - 2,3 - SA, limita a transmissão humano – aviária, exceção: H5N1; Porcos = dois receptores (local de recombinação – vírus potencialmente pandêmico), ex. H1N1 SOIV (pneumonia viral); Fatores ambientais , virais e do hospedeiro influenciam na aptidão e transmissão do Influenza. Aumento da replicação, transmissão e patogênese além de fatores adicionais modulam o potencial viral.
 e α - 2,3 - SA (aviário), ex. H1 e H3; Aves = galactose presente nos receptores α - 2,3 - SA, limita a transmissão humano – aviária, exceção: H5N1; Porcos = dois receptores (local de recombinação – vírus potencialmente pandêmico), ex. H1N1 SOIV (pneumonia viral); Fatores ambientais , virais e do hospedeiro influenciam na aptidão e transmissão do Influenza. Aumento da replicação, transmissão e patogênese além de fatores adicionais modulam o potencial viral.")
19
Competência de replicação
Adaptações na polimerase e hemaglutinina são importantes para o sucesso viral. Aptidão viral é crucial para o crescimento do vírus, onde adaptações aumentam a eficiência de replicação, ex. resíduo K 627 – confere habilidade de H1N1 e H3N2 infectarem humanos, enriquecimento da virulência de H5N1 e H7N7; D701N – confere adaptação e transmissão de AVI; R591 – confere eficiente replicação ao H1N1 mesmo sem os resíduos anteriores. Fatores do hospedeiro que influenciam a infeccção do vírus Influenza Entender a interação entre célula e vírus e desenvolvimento de novas pesquisas antivirais além de intervenção farmacológica. Fatores hospedeiros necessários: fusão viral, transporte de complexos virais do núcleo e para o núcleo, transcrição e tradução do genoma viral, montagem e saída do vírus. Ex. ATPase envolvida na endocitose viral, sensor de cálcio regulador de processos celulares, cinases, p27 (regulador celular). Interações entre as vias: deleção de componentes da via de Wnt aumentam a proliferação e diminuem a produção de interferon.
. Interações entre as vias: deleção de componentes da via de Wnt aumentam a proliferação e diminuem a produção de interferon.")
20
PATOGÊNESE VIRAL • Imunidade pré-existente (adquirida)
• Determinantes Virais • Determinantes do hospedeiro Fatores envolvidos na interação vírus-hospedeiro que contribuem para patogênese:

21
É possível que idosos possam ter algum nível de proteção cruzada
PATOGÊNESE VIRAL Imunidade pré-existente: É possível que idosos possam ter algum nível de proteção cruzada (cross-reactive antibodies) contra vírus Influenza H1N1 proveniente de anticorpos pré-existentes de outros vírus influenza → Prévia exposição a vírus antigeneticamente similares a H1N1
 contra vírus Influenza H1N1 proveniente de anticorpos pré-existentes de outros vírus influenza. → Prévia. exposição a vírus antigeneticamente similares a H1N1.")
22
PATOGÊNESE VIRAL ↓ Imunidade pré-existente:
Imunização com H1N1 de 1918, H1N1 humanos antes de 1947 ou H1N1 clássico de suínos mostraram a formação de reação cruzada de anticorpos ↓ Proteção ao vírus H1N1 de 2009 (próxima similaridade antigênica entre os vírus H1N1 de 1918 e de 2009)
")
23
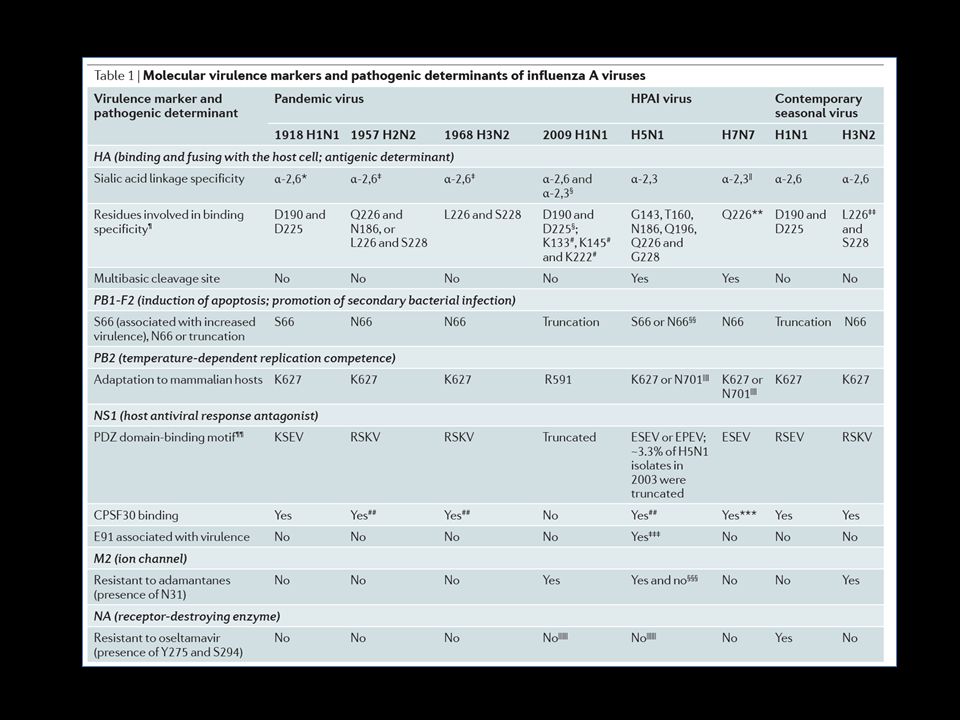
PATOGÊNESE VIRAL Determinantes virais:
PB1-F2 : proteína não estrutural pró-apoptótica; localiza-se na mitocôndria → apoptose; inibe resposta de IFN HA: hemaglutinina NS1: proteína não estrutural 1; antagonista de resposta antiviral (IFN) do hospedeiro A sequência do sítio de clivagem de HA é o maior determinante da gravidade da doença
 do hospedeiro. A sequência do sítio de clivagem de HA é o maior determinante da gravidade da doença.")
24
PATOGÊNESE VIRAL Determinantes virais:
NA: enzima neuraminidase destruidora de ácido siálico PB1, PB2 e PA: complexo da polimerase NP: nucleoproteína NS1: proteína não estrutural 1; antagonista de resposta antiviral (IFN) do hospedeiro e moduladora da resposta imune adaptativa
 do hospedeiro e moduladora da resposta imune adaptativa.")
25
Fatores determinantes da patogênese

26
Em humanos, o vírus da gripe aviária H5N1 pode gerar aumento de IFN no sangue induzido por proteínas como as interleucinas e citocinas. Estas se correlacionam com a carga viral elevada e leva a um aumento da frequência de morte.

27
Os vírus H5N1 e H1N1 geram uma resposta imune do hospedeiro maior e mais rápida do que outros vírus. Porém esta reposta gera a indução de macrófagos e infiltração de neutrófilos para ambas as linhagens(H5N1 e H1N1), que resulta numa inflamação pulmonar aguda e na desregulamentação de respostas antivirais.
, que resulta numa inflamação pulmonar aguda e na desregulamentação de respostas antivirais..")
28
Estudos com ratos e camundongos KO a algumas interleucinas provaram que há uma resistência maior destes aos vírus H1N1 e H5N1 Outros ratos KO não só a interleucinas mas também a quimiocina e TNF tiveram um tempo de vida similar a ratos WT. Isso demonstra que a eliminação de apenas alguns fatores pró-inflamatórios são vantajosos para o hospedeiro.

29
Porém outro estudo mostrou que a produção interleucina-10 poderia controlar a inflamação pulmonar e prováveis lesões produzidas pelo vírus.

30
Qual o futuro destas doenças?

31
Os vírus de gripe sazonal mudam com o tempo devido a uma seleção imune, isso faz com que hajam mudanças nas características antigênicas. Drogas antivirais como o Tamiflu e Relenza inibem a atividade da proteína NA, bloqueando assim a liberação dos vírions recém-formados das células infectadas. Essas drogas são eficazes mas com o tempo podem acabar gerando vírus mais resistentes.

32
Soluções Pesquisar novas formas terapêuticas de antivirais que ajam combatendo a infecção de inúmeras formas, sendo estas mais especificas para combater o vírus sem torná-lo mais resistente; Pesquisar para entendermos melhor o papel da IL-10 nos humanos; Procurar entender melhor a relação vírus com o hospedeiro e quais as vantagens que o hospedeiro pode trazer para o vírus.

33
PATOGÊNESE VIRAL Determinantes do hospedeiro:

Apresentações semelhantes







>")


.>")
, é transmitida de pessoa a pessoa, através da água e de alimentos contaminados com matéria fecal .EM.>")







