Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Capítulo 8 8.1 – Introdução 8.2 – Passagem através da membrana requere um aparato especial 8.3 – Translocação proteica pode ser pós-tradução ou co-tradução 8.4 – Chaperones podem ser requisitados para folding proteico 8.5 – Chaperones podem ser necessários pelas proteínas recém-sintetizadas e desnaturadas 8.6 – A família da Hsp70 é ubíqua

2
Figura 8.1: Mapeamento da célula em termos dos possíveis destinos finais das proteínas recém-sintetizadas e dos sistemas que as transportam.

3
Proteínas Associadas a membranas Sintetizadas no citosol
Não associadas a membranas Proteínas Sintetizadas em organelos (mitocôndrias, cloroplastos)
")
4
Capítulo 8 8.1 – Introdução 8.2 – Passagem através da membrana requere um aparato especial 8.3 – Translocação proteica pode ser pós-tradução ou co-tradução 8.4 – Chaperones podem ser requisitados para folding proteico 8.5 – Chaperones podem ser necessários pelas proteínas recém-sintetizadas e desnaturadas 8.6 – A família da Hsp70 é ubíqua

5
Translocação Proteica – processo de inserção ou passagem por uma membrana
Proteínas são transportadas para os peroxissomas por uma proteína de transporte Proteína Proteínas entram no RE ou nas mitocôndrias ou cloroplastos ligando-se a um transloncon que as atravessa pela membrana Proteína Transportador

6
As proteínas entram no núcleo passando por poros nucleares muito grandes. Embora forneça o ambiente que permite ao substrato entrar ou sair, não fornece o aparato que se liga so substrato e o desloca. Proteína Transportador

7
Capítulo 8 8.1 – Introdução 8.2 – Passagem através da membrana requere um aparato especial 8.3 – Translocação proteica pode ser pós-tradução ou co-tradução 8.4 – Chaperones podem ser requisitados para folding proteico 8.5 – Chaperones podem ser necessários pelas proteínas recém-sintetizadas e desnaturadas 8.6 – A família da Hsp70 é ubíqua

8
Existem duas formas da proteína fazer o contacto inicial com uma membrana:
A proteína “nascente” associa-se ao aparato de translocação enquanto está a ser sintetizada no ribossoma – Translocação Co-Tradução A proteína é liberada do ribossoma, após tradução, e difunde-se até à membrana apropriada onde se associa com o aparato de transporte – Translocação Pós-Tradução

9
Localização dos ribossomas
Ligados à membrana: Translocação Co-Tradução é usada para proteínas que entram no RE. Consequentemente, o ribossoma localiza-se à superfície do RE. Ribossomas Livres: Todos os outros ribossomas estão localizados no citosol. O destino das proteínas sintetizadas é variado: transportadas para o núcleo; associadas a organelos através de translocação pós-tradução.

10
Figura 8.5: Para se associar a uma membrana, a proteína necessita um sinal adequado, tipicamente um motivo (sequência)
")
11
Figura 8.6: O sinal para a translocação co-tradução localiza-se no terminal N, permitindo haver um reconhecimento do aparato de translocação. Isto acontece enquanto a proteína está a ser sintetizada.

12
Capítulo 8 8.1 – Introdução 8.2 – Passagem através da membrana requere um aparato especial 8.3 – Translocação proteica pode ser pós-tradução ou co-tradução 8.4 – Chaperones podem ser requisitados para folding proteico 8.5 – Chaperones podem ser necessários pelas proteínas recém-sintetizadas e desnaturadas 8.6 – A família da Hsp70 é ubíqua

13
Capazes de adquirir a sua conformação madura espontaneamente (auto-assemblagem)
Proteínas Aquisição de estrutura correcta necessita ajuda de chaperones

14
Figura 8.9: Folding proteico dá-se por interacções entre superfícies reactivas, tipicamente cadeias laterais hidrofóbicas. Se o processo não for controlado poderão ocorrer interacções incorrectas, que terão repercussões na funcionalidade e viabilidade da proteína. Chaperones controlam este processo.

15
Chaperones são proteínas que controlam a assemblagem correcta fazendo com que a proteína alvo adquira uma possível conformação ao invés de outras. Funcionam impedindo a formação de estruturas incorrectas

16
Capítulo 8 8.1 – Introdução 8.2 – Passagem através da membrana requere um aparato especial 8.3 – Translocação proteica pode ser pós-tradução ou co-tradução 8.4 – Chaperones podem ser requisitados para folding proteico 8.5 – Chaperones podem ser necessários pelas proteínas recém-sintetizadas e desnaturadas 8.6 – A família da Hsp70 é ubíqua

17
Ao sair do ribossoma a proteína encontra-se numa forma unfolded
Ao sair do ribossoma a proteína encontra-se numa forma unfolded. Os chaperones irão influenciar o processo de folding espontâneo controlando a acessibilidade às superfícies reactivas. Chaperones Quando uma proteína é desnaturada, há novas regiões expostas que podem interagir. Chaperones são capazes de detectar isto e vão assistir na renaturação ou na sua remoção por degradação.

18
Há dois grandes tipos de chaperones bem caracterizados:
Sistema Hsp70 – consiste em proteínas individuais que se ligam, e actuam sobre, os substratos cujo folding deve ser controlado. Os componentes deste sistema são: Hsp70, Hsp40 e GrpE. Sistema Chaperonin – consiste numa larga estrutura oligomérica. As proteínas unfolded são inseridas lá dentro onde o ambiente protegido direcciona o folding. GroEL/GroES encontram-se em todas as classes de organismos. TRiC só é encontrado no citosol eucariótico.

19
Figura 8.12: Sistemas e seus componentes

20
Capítulo 8 8.6 – A família da Hsp70 é ubíqua 8.1 – Introdução
8.2 – Passagem através da membrana requere um mecanismo especial 8.3 – Translocação proteica pode ser pós-tradução ou co-tradução 8.4 – Chaperones podem ser requisitados para folding proteico 8.5 – Chaperones podem ser necessários pelas proteínas recém-sintetizadas e desnaturadas 8.6 – A família da Hsp70 é ubíqua

21
Legenda: DnaJ (Hsp40); DnaK (Hsp70)
Figura 8.13: Hsp40 liga-se ao substrato e depois o Hsp70. A hidrólise de ATP leva a uma modificação conformacional. GrpE desloca o ADP o que leva à libertação dos chaperones. Podem ocorrer múltiplos ciclos. Esta família é encontrada em bactérias, citosol eucariótico, RE, cloroplastos e mitocôndrias

22
A Hsp70 reconhece motivos hidrofóbicos na proteína
A Hsp70 reconhece motivos hidrofóbicos na proteína. Numa proteína madura, bem estruturada, estes motivos fazem parte do interior (núcleo) hidrofóbico proteico. A exposição destes motivos, seja por desnaturação ou pelo facto de a proteína estar a ser sintetizada vai induzir a acção protectora dos sistemas de chaperones.
 hidrofóbico proteico. A exposição destes motivos, seja por desnaturação ou pelo facto de a proteína estar a ser sintetizada vai induzir a acção protectora dos sistemas de chaperones.")
23
Capítulo 8 8.1 – Introdução 8.2 – Passagem através da membrana requere um aparato especial 8.3 – Translocação proteica pode ser pós-tradução ou co-tradução 8.4 – Chaperones podem ser requisitados para folding proteico 8.5 – Chaperones podem ser necessários pelas proteínas recém-sintetizadas e desnaturadas 8.6 – A família da Hsp70 é ubíqua

24
8.7 – Hsp60/GroEL forma uma estrutura oligomérica em anel
A proteína alvo é levada para o interior de uma estrutura cilíndrica para folding ou degradação desta, sendo um processo controlado impossibilitando interações com outras proteínas.

25
Grande aparato formado por duas
subunidades: Hsp60 (GroEL) 14 subunidades arranjadas em dois anéis heptaméricos invertidos, constituindo um cilindro com duas metades iguais.
 14 subunidades. arranjadas em dois anéis heptaméricos. invertidos, constituindo um cilindro com. duas metades iguais.")
26
Hsp10 (GroES) um heptâmero
formando uma cúpula oca que se associa a uma das extremidades do cilindro, como uma tampa. GroEL: chaperonina (papel essencial) GroES: co-chaperonina
 GroES: co-chaperonina.")
27
Interações hidrofóbicas entre o substrato e resíduos de GroEL que estão expostos no interior da cavidade. Substrato e ATP se ligam ao mesmo anel de GroEL (anel proximal). Ligação de GroES ao anel proximal, induzindo mudança de conformação deste. Resíduos hidrofóbicos se ocupam da ligação de GroES. Substrato encontra-se em um meio hidrofílico, promovendo mudança na conformação do substrato. Hidrólise de ATP promove a energia necessária. GroES e substrato são liberados. Vários ciclos ocorrem até que a proteína alcance a conformação madura.
. Ligação de GroES ao anel proximal, induzindo mudança de conformação deste. Resíduos hidrofóbicos se ocupam da ligação de GroES. Substrato encontra-se em um meio hidrofílico, promovendo mudança na conformação do substrato. Hidrólise de ATP promove a energia necessária. GroES e substrato são liberados. Vários ciclos ocorrem até que a proteína alcance a conformação madura.")
28
8.8 – Seqüências sinais iniciam translocação
Proteínas associam-se ao sistema ER somente no caso de co-tradução, ou seja, só enquanto estão sendo sintetizadas.

29
aminoácidos N-terminais.
A seqüência sinal da proteína substrato é responsável pela sua associação à membrana. Geralmente, uma seqüência de 15-30 aminoácidos N-terminais. Diferentes sinais são necessários para indicar diferentes destinos das proteínas.

30
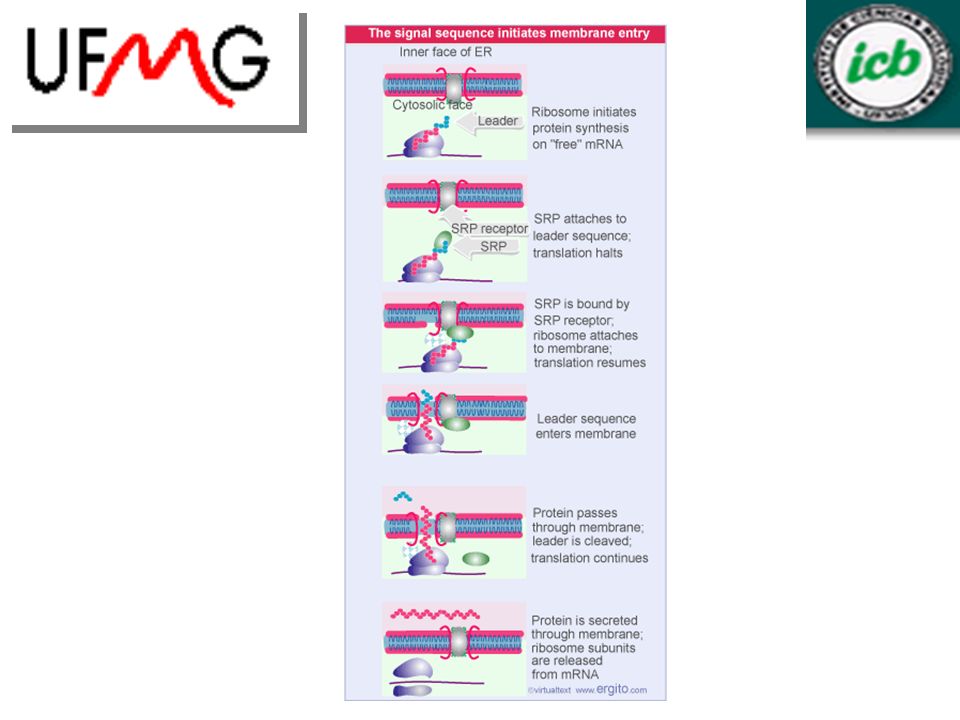
8.9 – A seqüência sinal interage com SRP
Translocação (estágios): Ribossomos carregando polipeptídios nascentes associam-se à membrana. Cadeia nascente é transferida para um canal e é translocada através dele. O agrupamento dos ribossomos à membrana requer uma partícula de reconhecimento de sinal (SRP), que se liga à seqüência sinal de uma proteína nascente secretada e à proteína localizada na membrana (receptor SRP).
: Ribossomos carregando polipeptídios nascentes associam-se à membrana. Cadeia nascente é transferida para um canal e é translocada através dele. O agrupamento dos ribossomos à membrana requer uma partícula de reconhecimento de sinal (SRP), que se liga à seqüência sinal de uma proteína nascente secretada e à proteína localizada na membrana (receptor SRP).")
32
8.10 – SRP interage com receptor SRP
SRP é um complexo de 7S RNA com 6 proteínas. Receptor SRP é um dímero contendo subunidades SRa e SRb: SRb é uma proteína de membrana integral enquanto a SRa possui uma porção que é ancorada pela SRb.

33
SRP carrega GDP quando se liga à seqüência sinal
SRP carrega GDP quando se liga à seqüência sinal. O ribossomo estimula a troca de GDP por GTP. Porém, a seqüência sinal inibe a hidrólise do GTP. Tanto SRP quanto receptor SRP realizam hidrólise de GTP quando a seqüência sinal é transferida para membrana.

34
8.11 – O translocon forma um poro
Translocon é o canal que atravessa a membrana do ER, formado por um complexo de proteínas residentes da membrana (Sec61a, b, g). - Canal aquoso, possibilita a passagem de proteínas hidrofílicas através da membrana. - O canal é fechado no interior do ER para que não haja passagem livre de íons.
. - Canal aquoso, possibilita a. passagem de proteínas hidrofílicas. através da membrana. - O canal é fechado no interior do. ER para que não haja passagem. livre de íons.")
35
- Uma proteína nascente é transferida
diretamente do ribossomo para o translocon. O ribossomo fecha o canal na extremidade do citosol.

36
8.12 – Translocação requer inserção no translocon e (às vezes) componentes adicionais são necessários Algumas proteínas nascentes requerem a presença de componentes adicionais, como por exemplo, TRAM que estimula a translocação de todas as proteínas.

37
Um aparato mais complexo é necessário quando ocorre uma translocação pós-tradução.
O complexo Sec61 forma o canal, mas outras quatro proteínas também são necessárias, além do chaperone BiP (membro da classe Hsp70) e de suplemento de ATP. BiP impede que a proteína que está sendo translocada volte para o citosol.
 e de. suplemento de ATP. BiP impede que a proteína que está. sendo translocada volte para o citosol.")
38
8.13 – Translocação reversa manda proteínas para o citosol para degradação
O ER funciona como um sistema de “controle de qualidade” em que as proteínas que não se encontram em suas conformações adequadas são identificadas e degradadas. Porém, a degradação não ocorre no ER e sim no citosol. Assim, tais proteínas devem ser exportadas de volta para o citosol através da translocação reversa.

39
O translocon Sec61 é também utilizado, mas não há associação com o ribossomo.
Não se sabe ainda qual o mecanismo de abertura do canal para que a proteína possa atravessar a membrana do ER até o citosol. - Uma possibilidade é que a proteína seja reconhecida por chaperones que a transferem para o translocon.

40
8.14 – Proteins reside in membranes by means of hydrophobic regions
Definição: as proteínas de membrana são aquelas que precisam do rompimento da membrana para serem liberadas Os resíduos interno e externo dessas proteínas podem variar de tamanho As proteínas com um único domínio transmembrana podem ser divididas em duas classes: Grupo I – o N-terminal da proteína é extracelular Grupo II – o N-terminal é intracelular LGCM URLGA

41
LGCM URLGA

42
Proteínas transmembranas podem também contar vários domínios
Proteínas transmembranas podem também contar vários domínios. Se esses forem em número par, o C e N-terminais serão no mesmo compartimento, se forem ímpar, estarão em áreas distintas LGCM URLGA

43
Os domínios transmembrana de proteínas com apenas um domínio não exercem funções importantes.
Proteínas com múltiplos domínios, por sua vez, tem em sua região transmembrana papeis importantes. URLGA

44
8.15 – Anchor sequences determine protein orientation
O que distingue uma proteína de membrana de outras? Seqüência sinal (tipo I e II) Seqüência “stop-transfer” aminoácidos hidrofóbicos, resíduos iônicos Na inserção das proteínas do tipo I, a seqüência sinal no N-terminal. A seqüência stop-transfer está no C-terminal.
 Seqüência stop-transfer aminoácidos hidrofóbicos, resíduos iônicos. Na inserção das proteínas do tipo I, a seqüência sinal no N-terminal. A seqüência stop-transfer está no C-terminal.")
45
No caso de proteínas tipo II, a seqüência sinal e a seqüência de ancoragem (anchor sequence) estão combinadas.
 estão combinadas.")
46
A composição de aa é mais importante que a seqüência
As extremidades são positivas e o interior é neutro As cargas positivas são localizadas no citoplasma

47
8.16 – How do proteins insert into membranes?
Ainda não é conhecido o mecanismo pelo qual as proteínas de membrana sai do canal e entra na bicamada lípídica Uma das possibilidades sugeridas (figura abaixo) é que existiria um mecanismo próprio de transferência direta para a membrana. Esse mecanismo é sempre determinado por uma seqüência transmembrana no poro.
 é que existiria um mecanismo próprio de transferência direta para a membrana. Esse mecanismo é sempre determinado por uma seqüência transmembrana no poro.")
48
8.17 – Post-traslational membrane insertion depends on leader sequences
Mitocôndrias e cloroplastos possuem mecanismos pós-traducionais que reconhecem a membrana externa da organela (figura) A proteína passa pela membrana, e a seqüência sinal é clivada por proteases na organela.
 A proteína passa pela membrana, e a seqüência sinal é clivada por proteases na organela.")
49
A seqüência sinal possui aminoácidos básicos e hidrofóbicos.
O reconhecimento da seqüência não depende da seqüência em si, mas da capacidade de formar uma hélice que contenha um lado com aminoácidos hidrofóbicos e outro com aa básicos A seqüência sinal e o polipeptídeo se enovelam independentemente A seqüência deve ser capaz de formar uma estrutura capaz de ser reconhecida pelos receptores da organela O polipeptídio não desempenha nenhuma função no reconhecimento

50
8.18 – A hierarchy of sequences determines location within organelles
O caminho “default”das proteínas na mitocôndria é de se mover para a matriz Proteínas que devem se dirigir para o espaço intermembrana para a membrana interna devem possuir um sinal adicional A primeira parte do sinal indica qual é a organela de destino, e a segunda parte, qual é necessária se o destino não for a matriz A primeira clivagem é necessária para as proteínas que residem na matriz. Caso o destino seja outro, uma segunda clivagem do sinal é necessária. Ou seja, a proteína primeiro alcança a matriz para depois ir para o seu destino

51
A passagem pela membrana do cloroplasto é semelhante a da mitocôndria.

52
8.19 – Inner and outer mitochondrial membranes have different translocons
Receptores de proteínas em que a seqüência sinal é interna Subcomplexos com domínios expostos no citosol. Reconhecem o N-terminal das proteínas exportadas pra a mitocôndria Componentes do canal . São dois os subcomplexos que são receptores de superfície Canal de translocaçâo

53
Os receptores de transporte da membrana são chamados TOC e TIC no cloroplasto e TOM e TIM na mitocôndria, sendo que eles se referem a membrana externa e interna, respectivamente. Na mitocôndria, as proteínas são passadas diretamente do TOM para o TIM. São dois os complexos TIM na membrana interna: Tim17-13 que transporta proteínas para o lúmen e Tim22-54 que transporta proteínas para a membrana interna. Na matriz, Tim17-23 se associa a Tim44, que por sua vez se associa as chaperonas Hsp70 e MGE Essa associação garante que a proteína não-novelada essa “puxada” corretamente pelo canal.

54
Para as proteínas acharem seu caminho do TOM para o TIM, elas são escoltadas por Tim9-10 e Tim8-13
Tim9-10 pode dirigir o substrato tanto para Tim17-23 e 22-54 Tim8-13 dirige para apenas Tim22-54

55
Uma proteína pode passar diretamente por TOM Tim9-10 Tim22-54
É possível que uma proteína só tenha sua conformação correta na mitocôndria.

56
8.20 – Peroxisomes employ another type of translocation system
Todos os componentes dos peroxissomos são importados do citosol O transporte de proteínas é pós-traducional As proteínas importadas para a matriz possuem seqüências sinal pequenas, chamadas PTS1 e PTS2 Os receptores do peroxissomos são o Pex5p e Pex7p As proteínas são importadas já enoveladas

57
Pex5p e Pex7p não são transmembrana, mas possuem uma grande região no citosol
Elas ciclam as proteínas do citosol para o interior da organela

58
8.21. Bactérias usam translocação co e pos-translacional
Capítulo 8 – Genes VIII 8.21. Bactérias usam translocação co e pos-translacional 8.22 Sistema Sec transporta proteínas para e através da membrana interna 8.23 Sistema de tradução na E.coli independente do sistema Sec 8.24 Poros são usados para importação e exportação nuclear 8.25 Estrutura simétrica dos poros 8.26 O poro nuclear é uma estrututa tamanho dependente para substâncias pequenas Ruth Agostinho. Abril, 2005

59
8.21 Bactérias usam translocação co e pos-translacional
Proteínas produzidas no citoplasma podem manter-se no interior da célula ou ser secretadas. Proteínas exportadas seguem vários destinos: Periplasma Memb. externa Memb. interna Fig.1: Destino das proteínas bacterianas. é inserida na membrana interna; atravessa a membrana interna e vai para o periplasma; é inserida na membrana externa; atravessa a membrana externa e vai para o meio.

60
8.21 Bactérias usam translocação co e pos-translacional
Este transporte de proteínas é levado a cabo por diferentes complexos proteicos da membrana interna, dependendo se vão dentro da membrana ou atravessá-la. -A importação é feita tanto co como pos-translacionalmente. -A secreção de algumas proteínas pode feita co como pos-translacionalmente. As proteínas bacterianas exportadas possuem sequências N-terminal líderes com uma terminação N hidrofílica e um core hidrofóbico adjacente. Mutações neste N-terminal impedem a secreção das mesmas. Estes genes são então suprimidos por mutações em outros genes, componentes do aparato de exportação proteico.

61
8.22 Sistema Sec transporta proteínas para e através da membrana interna
Sistema Sec é o mais bem caracterizado sistema de transporte de proteínas através da membrana. Fig.2: Sistema Sec. Possui o translocon SecYEG embebido na membrana, a proteína associada SecA (empurra a proteína a ser transferida pelo canal), o chaperone SecB (transfere proteínas nascentes para o SecA) e a peptidase sinal (cliva as sequências N-terminais das protéinas translocadas).
, o chaperone SecB (transfere proteínas nascentes para o SecA) e a peptidase sinal (cliva as sequências N-terminais das protéinas translocadas).")
62
8.22 Sistema Sec transporta proteínas para e através da membrana interna
Há duas maneiras de predominantes de dirigir uma proteína para o canal Sec: - associá-la ao chaperone Sec B; - associá-la à base 4.5S SecB é o menos abundante exercendo, no entanto, duas funções bastante importantes na promoção da exportação de proténas: - comporta-se como um chaperone ligando-se a uma proteína nascente impedindo foldings indesejados; - Tem afinidade com o SecA o que lhe permite ligar percursores de proteínas à membrana.

63
8.22 Sistema Sec transporta proteínas para e através da membrana interna
SecA é uma proteína membranar periférica com formas alternativas de se ligar à membrana: -como proteína periférica associa-se à membrana devido à sua afinidade aos lípidos ácidos e aos SecY componente do translocon -na presença de outras proteínas (SecD e SecF) compota-se como uma proteína de spanning, ou seja, como um motor que “puxa” a proteína do substrato através do translocon SecYEG.
 compota-se como uma proteína de spanning, ou seja, como um motor que puxa a proteína do substrato através do translocon SecYEG.")
64
SecA tem também uma actividade de ATPase:
8.22 Sistema Sec transporta proteínas para e através da membrana interna SecA tem também uma actividade de ATPase: 1. Liga-se a um percursor de proteína (transferida pelo SecB), 2. Liga-se a uma molécula de ATP, 3. ~20 aa são translocados através da membrana, 4. Hidrólise do ATP percursor liberta-se do SecA. Fig.3: SecA insere a proteína no canal de membrana, translocação que requere a hidrólise do ATP e sucessivos associações e desassociações com o canal.
, 2. Liga-se a uma molécula de ATP, 3. ~20 aa são translocados através da membrana, 4. Hidrólise do ATP. percursor liberta-se do SecA. Fig.3: SecA insere a proteína no canal de membrana, translocação que requere a hidrólise do ATP e sucessivos associações e desassociações com o canal.")
65
8.22 Sistema Sec transporta proteínas para e através da membrana interna
Uma vez liberto do SecA, o percursor pode atravessar a membrana devido à diferença de potencial eléctrico ao longo da membrana. Este mecanismo de transferência de percursores por diferença de potencial é incapaz de iniciar a transferência sozinho, mas uma vez iniciado o ciclo pela SecA ATPase, este é capaz de o continuar.

66
8.23 Sistema de tradução na E.coli independente do sistema Sec
E.coli e organelos têm sistemas de translocação de proteínas muito relacionados. E. coli desenvolveu um sistema alternativo que permite que certas proteínas sejam inseridas na membrana sem o auxílio de um aparato de translocação. Fig.4: Contacto inicial electrostático. - Regiões N e C-terminais da proteína interagem com as cabeças negativas dos fosfolípidos.

67
8.23 Sistema de tradução na E.coli independente do sistema Sec
Fig.5: Líder hidrofóbico inserido. - A proteína penetra na membrana usando os grupos hidrofóbicos no N-terminal e uma sequência âncora interna. Fig.6: Proteína translocada. 2 1 Força - A hidrofobicidade (1) é a força principal para a translocação, embora possa haver uma outra força auxiliar (2) entre o periplasma positivo e a região ácida da proteína.
 é a força principal para a translocação, embora possa haver uma outra força auxiliar (2) entre o periplasma positivo e a região ácida da proteína.")
68
8.24 Poros são usados para importação e exportação nuclear
O transporte entre o núcleo e o citoplasma dá-se nos dois sentidos, do núcleo para o citolplasma e deste para o núcleo. Proteínas que são produzidas no citosol e são transportadas deste para o núcleo. RNA e proteínas sintetizados no núcleo e transportados para o citosol. Fig.7: Substâncias importadas e exportadas pelo núcleo.

69
8.25 Estrutura simétrica dos poros
Fig.8: Poros nucleares ao microscópio electrónico . Estruturas anelares. Fig.9: Modelo a 3D de um poro nuclear. 8 sub-unidades.

70
8.25 Estrutura simétrica dos poros
Fig.10: Componentes do poro nuclear. Anéis de ~120nm 8 braços radiais responsáveis pelo ancoramento das unidades ao invólucro nuclear 8 sub-unidades interiores que fecham o poro e reduzem o seu diâmetro para ~48nm. Dentro deste espaço está o transportador com uma abertura de ~10nm. Cada unidade é constituída por ~30 partículas diferentes, proteínas estas encontradas nos dois lados do poro.

71
8.26 O poro nuclear é uma estrututa tamanho dependente para substâncias pequenas
A capacidade do material de difundir livremente pelos poros nucleares está limitada pelo seu tamanho. Membrana nuclear permeável a moléculas menores que 5000D (iões, nucleotídeos) Proteínas entre 5 e 50kD difundem a uma velocidade inversamente proporcional ao seu tamanho por difusão passiva. Proteínas maiores que 50kD entram no núcleo por transporte activo. À medida que se aumenta o tamanho do material, a passagem pelo núcleo vai diminuindo. Proteínas maiores obrigam o núcleo a aumentar a sua área de passagem. O núcleo tem um mecanismo de abertura que permite que o seu interior se expanda até ~20nm, à medida que o material o atravessa.
 Proteínas entre 5 e 50kD difundem a uma velocidade inversamente proporcional ao seu tamanho por difusão passiva. Proteínas maiores que 50kD entram no núcleo por transporte activo. À medida que se aumenta o tamanho do material, a passagem pelo núcleo vai diminuindo. Proteínas maiores obrigam o núcleo a aumentar a sua área de passagem. O núcleo tem um mecanismo de abertura que permite que o seu interior se expanda até ~20nm, à medida que o material o atravessa.")
72
Proteínas requerem sinais para serem transportadas através do poro.
Motivo de importação mais comum: NLS Não conservado; Forma, basicidade Resíduo de prolina que impede a formação de uma alfa-hélice, resíduos hidrofóbicos raros; Bipartidos Reconhecidos pelo mesmo sistema de transporte. Motivo de exportação: NES sequencia de ~ 10 aminoácidos; Padrão de leucinas conservado Uma proteína pode ter tanto um NLS quanto um NES Podem operar constitutivamente ou de forma regulada. URLGA

73
Receptores carreiam as proteínas transportadas pelo poro.
Uma proteína carreadora transporta o substrato através do poro e depois deve retornar para um novo ciclo; Importinas e Exportinas; Vias múltiplas de transporte: Pelo menos 2 diferentes na importação de proteínas; Cada classe de RNA possui um sistema diferente de exportação; Cada transportador reconhece uma seqüência diferente em seu substrato: especificidade. O transporte pode ser dividido em duas etapas: ancoragem e translocação. Para a ancoragem é necessário uma fração proteica; Apenas na presença de ATP uma proteína pode ser translocada. Isso envolve uma diferente fração da proteína. O receptor é a peça alvo no processo de ancoragem: liga-se ao poro e ao substrato: monômeros ou dímeros

75
Ran controla a direção do transporte
O poro nuclear não possui nenhuma proteína com atividade motora: Ran Proteína G monomérica em duas formas: Ran-GTP, no núcleo (Ran-GEF) – exportação; Ran-GDP, no citoplasma (Ran-GAP) – importação. Ex.: Importina-ab, NLS-proteína – estável com Ran-GDP - Formação estável apenas no citoplasma - Ran-GTP: dissociação do complexo ab no núcleo e liberação do substrato. Após a liberação de um importina no núcleo, ela vira substrato para uma exportina; O transporte através do poro não exige energia: importinas e exportinas se movem pelo poro por difusão facilitada (?). Interação com nucleoporinas. O grau de interação dos diferentes transportadores com as nucleoporinas permitem o transporte em diferentes velocidades. O direcionamento do transporte durante a passagem através do poro ainda é desconhecido.
 – exportação; Ran-GDP, no citoplasma (Ran-GAP) – importação. Ex.: Importina-ab, NLS-proteína – estável com Ran-GDP. - Formação estável apenas no citoplasma. - Ran-GTP: dissociação do complexo ab no núcleo e. liberação do substrato. Após a liberação de um importina no núcleo, ela vira substrato para uma exportina; O transporte através do poro não exige energia: importinas e exportinas se movem pelo poro por difusão facilitada ( ). Interação com nucleoporinas. O grau de interação dos diferentes transportadores com as nucleoporinas permitem o transporte em diferentes velocidades. O direcionamento do transporte durante a passagem através do poro ainda é desconhecido.")
76
RNA é exportado por diversos sistemas
O maiores substratos para exortação do núcleo são ribonucleoproteínas – ribossomos, mRNAs, snRNAs e tRNAs complexados a proteínas 3 primeiros: uma das proteínas do complexo exerce o papel de trabsportador. tRNAs ligam-se diretamente a uma proteína de exportação específica. Algumas proteínas vão e voltam ao núcleo constantemente: proteínas ligadas a poly(A)-RNA – motivo responsável funciona como sinal os dois transportes. A distinção de um RNA processado de precursores não processados está relacionado ao tempo: Interação do transportador com proteínas do aparato de transcrição. Splicing deve terminar. Mex67 ou TAP: interação com nucleoporinas
-RNA – motivo responsável funciona como sinal os dois transportes. A distinção de um RNA processado de precursores não processados está relacionado ao tempo: Interação do transportador com proteínas do aparato de transcrição. Splicing deve terminar. Mex67 ou TAP: interação com nucleoporinas.")
77
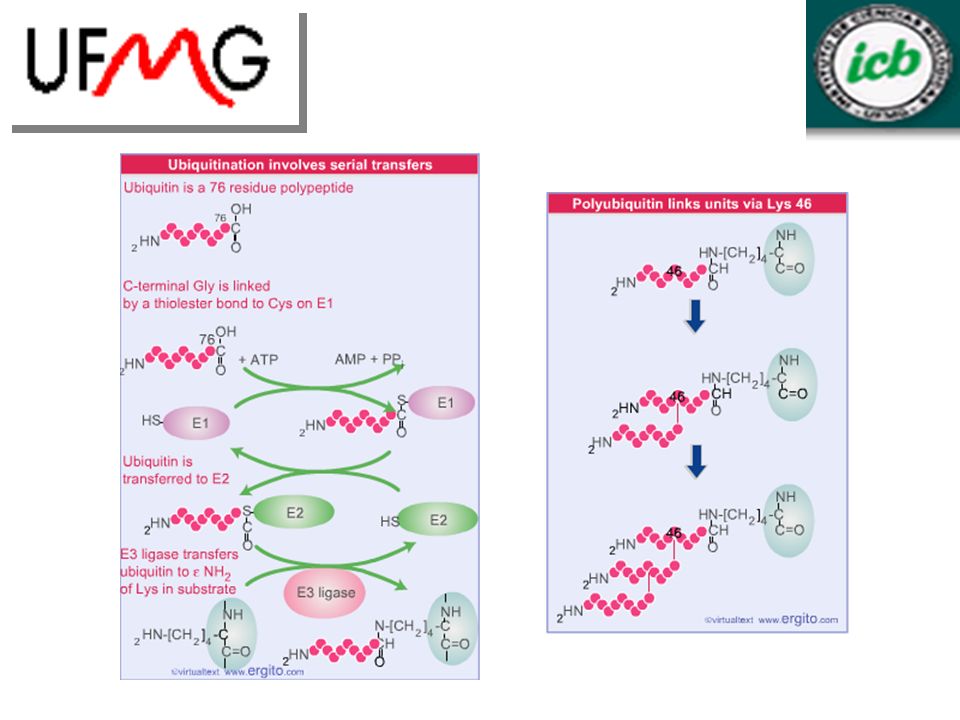
Ubiquitinação marca proteínas para degradação
Degradação proteíca: marcação e proteólise. Marcação: Adição covalente de Ubiquitina – mediado por 3 componentes: Enzima ativadora de ubiquitina, E1, que se liga à ubiquitina com gasto de ATP; Transferência da ubiquitina à enzima conjugada a ubiquitina, E2; Ubiquitina ligase, E3, transfere a ubiquitina ao substrato através de uma ligação peptídica a um resíduo de lisina. A escolha dos substratos para degradação é feita por E3 e E2. Vários resíduos de ubiquitina devem ser adicionados ao substrato para que esta possa ser degradada pelo proteasoma. A ubiquitinação pode ser revertida por proteases que clivam o conjugado de ubiquitina.

79
O proteasoma é grande máquina de degradação de proteínas ubquitinadas
Uma célula contém diversas proteases com especificidades variadas Proteases envolvidas no processamento de proteínas para que estas adquiram sua forma madura; Proteases do lisossoma, que degradam proteínas importadas para a célula O proteassoma, um grande complexo que degrada proteínas citoplasmáticas. Está envolvido tanto em degradação geral e eventos específicos de processamento (proteínas mal formadas). O proteassoma existe em duas formas: Um complexo 20S com atividade de protease. Proteínas adicionais convertem o complexo para a forma 26S. Tais unidades exercem atividade regulatória, conferindo especificidade. Tal conversão exige quebra de ATP.
. O proteassoma existe em duas formas: Um complexo 20S com atividade de protease. Proteínas adicionais convertem o complexo para a forma 26S. Tais unidades exercem atividade regulatória, conferindo especificidade. Tal conversão exige quebra de ATP.")
80
O proteasoma é grande máquina de degradação de proteínas ubquitinadas
O complexo 20S possui uma forma cilíndrica e os componentes adicionais se ligam ao final do cilindro. Os sítios proteolíticos estão no interior do cilindro e o acesso se da por uma fenda estreita – acesso de proteínas desnaturadas. O trabsporte e a hidrólise de proteínas marcadas exige a quebra de ATP. O mecanismo proteolítico do proteasoma envolve o ataque da ligação peptídica pelo grupo hidroxila de uma resíduo de treonina. O proteasoma possui proteases com diferentes especificidades - diferentes subunidades b. A degradação feita peloproteasoma é completa, sem que haja liberação de intermediários. Responsável por degradação em massa, degradação de novas proteínas mal formadas, clivagem de antígenos para apresentação via MHC e clivagem de proteínas relacionadas ao controle do ciclo celular.

Apresentações semelhantes



>")














>")

