Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Predição de Estrutura Secundária Marcilio Souto DIMAp/UFRN
Proteínas Predição de Estrutura Secundária Marcilio Souto DIMAp/UFRN

2
Somos seres protéicos A vida está intimamente ligada às proteínas
Estas moléculas especiais realizam as mais variadas funções no nosso organismo Transporte de nutrientes e metabólitos, catálise de reações biológicas Apesar da complexidade de suas funções, as proteínas são relativamente simples: Repetições de 20 unidades básicas, os aminoácidos

3
Aminoácido Um aminoácido consiste em um caborno “central” com uma ligação a grupo amino (-NH2), outra a um grupo carboxila (-COOH), a terceira a um átomo de hidrogênio e a quarta a uma cadeia lateral variável COO- | H3N+--C--H R
, outra a um grupo carboxila (-COOH), a terceira a um átomo de hidrogênio e a quarta a uma cadeia lateral variável. COO- | H3N+--C--H. R.")
4
Aminoácidos Single- & three-letter amino acid codes Additional codes
G Glycine Gly P Proline Pro A Alanine Ala V Valine Val L Leucine Leu I Isoleucine Ile M Methionine Met C Cysteine Cys F Phenylalanine Phe Y Tyrosine Tyr W Tryptophan Trp H Histidine His K Lysine Lys R Arginine Arg Q Glutamine Gln N Asparagine Asn E Glutamic Acid Glu D Aspartic Acid Asp S Serine Ser T Threonine Thr Additional codes B Asn/Asp Z Gln/Glu X Any amino acid

5
Definição As proteínas são macromoléculas complexas, compostas de aminoácidos, e necessárias para os processos químicos que ocorrem nos organismos vivos São os constituintes básicos da vida: tanto que seu nome deriva da palavra grega "proteios", que significa "em primeiro lugar” Nos animais, as proteínas correspondem a cerca de 80% do peso dos músculos desidratados, cerca de 70% da pele e 90% do sangue seco. Mesmo nos vegetais as proteínas estão presentes.

6
Importância A importância das proteínas, entretanto, está relacionada com suas funções no organismo, e não com sua quantidade Todas as enzimas conhecidas, por exemplo, são proteínas Muitas vezes, as enzimas existem em porções muito pequenas. Mesmo assim, estas substâncias catalisam todas as reações metabólicas e capacitam aos organismos a construção de outras moléculas - proteínas, ácidos nucléicos, carboidratos e lipídios - que são necessárias para a vida.

7
Polipeptídeos As proteínas também são chamadas de polipeptídeos, porque os aminoácidos que as compõe são unidos por ligações peptídicas Uma ligação peptídica é a união do grupo amino (-NH2) de um aminoácido com o grupo carboxila (-COOH) de outro aminoácido, através da formação de uma amida
 de um aminoácido com o grupo carboxila (-COOH) de outro aminoácido, através da formação de uma amida.")
8
Estrutura da Proteínas
Embora sejam quase inúmeras, todas as proteínas são formadas exclusivamente por apenas 20 aminoácidos, que se repetem numa seqüência característica para cada proteína Esta seqüência, conhecida como estrutura primária, é que, de fato, determina a forma e a função da proteína. A estrutura primária é somente a sequência dos amino ácidos, sem se preocupar com a orientação espacial da molécula As interações intermoleculares entre os aminoácidos das proteínas fazem com que a cadeia protéica assuma uma estrutura secundária e uma estrutura terciária.

9
Estrutura Secundária A estrutura secundária é uma função dos ângulos formados pelas ligações peptídicas que ligam os aminoácidos "The secondary structure of a segment of polypeptide chain is the local spatial arrangement of its main-chain atoms without regard to the conformation of its side chains or to its relationship with other segments". A conformação espacial é mantida graças as interações intermoleculares (ligação hidrogênio) entre os hidrogênios dos grupos amino e os átomos de oxigênio dos outros amino ácidos.
 entre os hidrogênios dos grupos amino e os átomos de oxigênio dos outros amino ácidos.")
10
Estrutura Secundária Em geral, estas ligações forçam a proteína a assumir uma forma helicoidal, como uma corda enrolada em torno de um tubo imaginário. Esta forma, a mais comum, é chamado de alfa hélice. Outras duas formas na estrutura secundária são as beta-sheets e turns. Nas beta-sheets, um segmento da cadeia interage com outro, paralelamente.

11
-Hélice É a forma mais comum de estrutura secundária regular
Caracteriza-se por uma hélice em espiral formada por 3,6 resíduos de aminoácidos por volta As cadeias laterais dos aminoácidos se distribuem para fora da hélice A principal força de estabilização da a - Hélice é a ponte de hidrogênio.

12
-Folhas Envolve 2 ou mais segmentos polipeptídicos da mesma molécula ou de moléculas diferentes, arranjados em paralelo ou no sentido anti-paralelo Os segmentos em folha da proteína adquirem um aspecto de uma folha de papel dobrada em pregas. As pontes de hidrogênio mais uma vez são a força de estabilização principal desta estrutura

13
Estrutura Terciária A estrutura terciária relaciona-se com os loopings e dobraduras da cadeia protéica sobre ela mesma. É a conformação espacial da proteína, como um todo, e não de determinados segmentos particulares da cadeia protéica. A forma das proteínas está relacionada com sua estrutura terciária. Existem, por exemplo, proteínas globulares (que tem forma esférica).
.")
14
Estrutura Terciária O que determina a estrutura terciária são as cadeias laterais dos aminoácidos Algumas cadeias são tão longas e hidrofóbicas que perturbam a estrutura secundária helicoidal, provocando a dobra ou looping da proteína. Muitas vezes, as partes hidrofóbicas da proteína agrupam-se no interior da proteína dobrada Longe da água e dos íons do ambiente onde a proteína se encontra, deixando as partes hidrofílicas expostas na superfície da estrutura da proteína. Regiões como "sítio ativos", "sítios regulatórios" e módulos são propriedades da estrutura terciária

15
Estrutura Terciária

16
Estrutura Quaternária
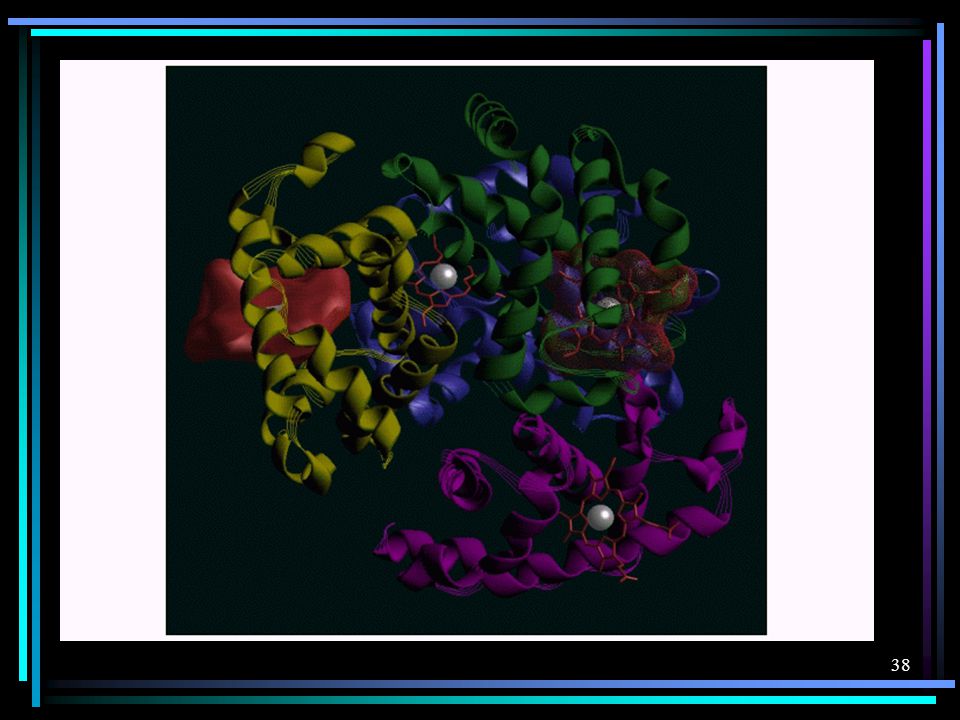
Existe, finalmente, a estrutura quaternária Ccertas proteínas, tal como a hemoglobina, são compostas por mais de uma unidade polipeptídica (cadeia protéica). A conformação espacial destas cadeias, juntas, é que determina a estrutura quaternária. Esta estrutura é mantida pelas mesmas forças que determinam as estruturas secundárias e terciárias. A figura ao lado mostra uma imumoglobulina que é, na verdade, um tetrâmero, isto é, constituída por 4 cadeias protéicas (polipeptídeos).
. A conformação espacial destas cadeias, juntas, é que determina a estrutura quaternária. Esta estrutura é mantida pelas mesmas forças que determinam as estruturas secundárias e terciárias. A figura ao lado mostra uma imumoglobulina que é, na verdade, um tetrâmero, isto é, constituída por 4 cadeias protéicas (polipeptídeos).")
17
Estrutura Quaternária
A figura ao lado mostra uma imumoglobulina que é, na verdade, um tetrâmero, isto é, constituída por 4 cadeias protéicas (polipeptídeos).
.")
18
Proteínas Conjugadas As proteínas podem ser simples
Constituidas somente por aminoácidos ou conjugadas Contêm grupos prostéticos, isto é, grupos não aminoácidos, tais como carbohidratos, íons, pigmentos, etc. A hemoglobina é um exemplo de proteína conjugada: contém 4 grupos prostéticos, cada um consistindo de um íon de ferro e a porfirina. São justamente estes grupos que habilitam a hemoglobina a carregar o oxigênio através da corrente sanguínea. As liproproteínas, tal como LDL e HDL, são também exemplos de proteínas conjugadas - neste caso, com lipídeos.

19
Proteínas Conjugadas

20
Outras Classificações
Uma outra forma de classificar as proteínas é baseado na sua função. Sobre este prisma, elas podem ser divididas em dois grupos: proteínas estruturais e proteínas biologicamente ativas Algumas proteínas, entretanto, podem pertencer aos dois grupos A maioria das proteínas estruturais são fibrosas - compostas por cadeias alongadas. Dois bons exemplos, nos animais, são o colágeno (ossos, tendões, pele e ligamentos) e a queratina (unhas, cabelos, penas e bicos).
 e a queratina (unhas, cabelos, penas e bicos).")
21
Outras Classificações
A grande maioria das proteínas biologicamente ativas são globulares, e sua atividade funcional é intrínsica a sua organização espacial Exemplos são as enzimas, hormônios protéicos (que atuam como mensageiros químicos), proteínas de transporte (como as lipo-proteínas, que podem carregar o colesterol) e imunoglobulinas (ou anticorpos), que protegem o corpo de microorganimos invasores. Muitas proteínas biologicamente ativas ficam na região da membrana celular, e atuam de diversas maneiras
, proteínas de transporte (como as lipo-proteínas, que podem carregar o colesterol) e imunoglobulinas (ou anticorpos), que protegem o corpo de microorganimos invasores. Muitas proteínas biologicamente ativas ficam na região da membrana celular, e atuam de diversas maneiras.")
22
Outras Classificações
A figura ao lado mostra uma porina, uma proteína trans-membrana, que atua como um canal iônico em bactérias. Existe um "buraco" na estrutura protéica, de cerca de 11 angstrons de diâmetro, onde os íons passam, seletivamente

23
Enzimas As enzimas são uma classe muito importante de proteínas biologicamente ativas. Elas são responsáveis pela catálise de diversas reações em nosso organismo. Reações que, sem o auxílio das enzimas, jamais aconteceriam ou, ainda, gerariam indesejados produtos colaterais. Em uma proteína enzimática, existe um certo domínio chamado de "sítio ativo", que liga-se ao substrato - a molécula reagente - e diminui a energia do estado de transição que leva ao produto desejado. A ligação entre o sítio ativo e o substrato é extremamente específica: a molécula precisa ter certas características eletrônicas e espaciais que permitam o seu "encaixe" com a proteína. Por isso esta relação tem sido chamada de lock'n'key, ou seja, chave-fechadura.

24
Enzimas: Sítio Ativo No exemplo da figura, uma determinada região da proteína liga-se à um substrato, que se adapta ao sítio ativo da enzima tal como uma chave faz a sua fechadura.

25
Enzimas: Inibidor A atividade de uma enzima pode ser bloqueada pela ação de outra molécula, um inibidor. Quando um inibidor interage com uma determinada região da enzima, chamado de sítio regulatório, provoca uma alteração na sua conformação e uma desativação do sítio catalítico. A atividade enzimática, portanto, pode ser controlada, pelo organismo, através da liberação ou captação de inibidores.

26
Enzimas: Inibidor

27
Caso tenham esquecido A sequência dos amino ácidos em todas as proteínas - fator que é responsável por sua estrutura e função - é determinado geneticamente a partir da sequência dos nucleotídeos no DNA celular. Quando uma proteína em particular é necessária, o código do DNA (gene) para esta proteína é transcrito em uma sequência complementar de nucleotídeos ao longo de um segmento de RNA - chamado de RNA mensageiro. Este segmento de RNA serve como uma forma para a síntese da proteína subsequente: cada grupo de 3 nuclueotídeos especifica um determinado aminoácido; estes aminoácidos são ligados na sequência codificada pelo RNA. No final do processo, obtém-se a proteína completa, cuja sequência de aminoácidos foi ditada pelo RNA mensageiro. Desta maneira, o organismo é capaz de sintetizar as várias proteínas com as funções mais diversas de que precisa.
 para esta proteína é transcrito em uma sequência complementar de nucleotídeos ao longo de um segmento de RNA - chamado de RNA mensageiro. Este segmento de RNA serve como uma forma para a síntese da proteína subsequente: cada grupo de 3 nuclueotídeos especifica um determinado aminoácido; estes aminoácidos são ligados na sequência codificada pelo RNA. No final do processo, obtém-se a proteína completa, cuja sequência de aminoácidos foi ditada pelo RNA mensageiro. Desta maneira, o organismo é capaz de sintetizar as várias proteínas com as funções mais diversas de que precisa.")
28
Previsão de Estrutura de Proteínas
Experimental Cristalização Raios X Ressonância nuclear magnética Cerca de 10 a 12 mil estruturas em repositórios públicos Processo caro e demorado Teórico Homologia Ab Inition Threading Aprendizado de Máquina

29
Modelagem por Homologia
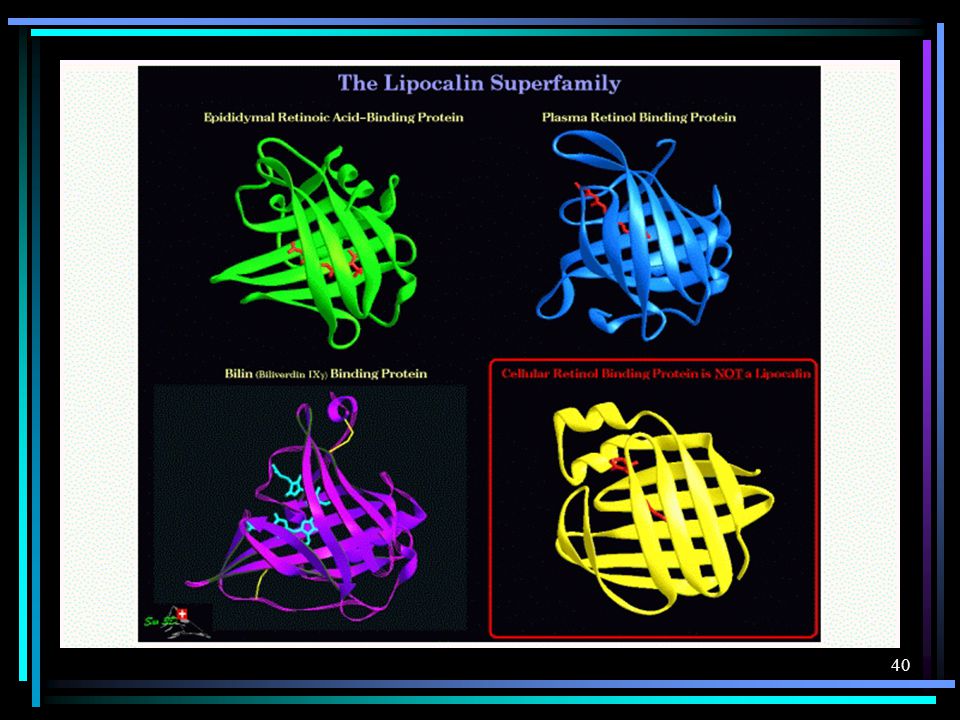
A ferramenta mais bem sucedida de predição de estruturas tridimensionais de proteínas é a modelagem por homologia, também conhecida como modelagem comparativa. Esta abordagem baseia-se em alguns padrões gerais que têm sido observados, em nível molecular, no processo de evolução biológica: homologia entre seqüências de aminoácidos implica em semelhança estrutural e funcional; proteínas homólogas apresentam regiões internas conservadas (principalmente constituídas de elementos de estrutura secundária: hélices-a e fitas-b); as principais diferenças estruturais entre proteínas homólogas ocorrem nas regiões externas, constituídas principalmente por alças ("loops"), que ligam os elementos de estruturas secundárias.
; as principais diferenças estruturais entre proteínas homólogas ocorrem nas regiões externas, constituídas principalmente por alças ( loops ), que ligam os elementos de estruturas secundárias.")
30
Modelagem por Homologia
Outro fato importante é que as proteínas agrupam-se em um número limitado de famílias tridimensionais. Estima-se que existam cerca de famílias protéicas. Conseqüentemente, quando se conhece a estrutura de pelo menos um representante de uma família, é geralmente possível modelar, por homologia, os demais membros da família.

31
Modelagem por Homologia
A modelagem de uma proteína (proteína-problema) pelo método da homologia baseia-se no conceito de evolução molecular. Isto é, parte-se do princípio de que a semelhança entre as estruturas primárias desta proteína e de proteínas homólogas de estruturas tridimensionais conhecidas (proteínas-molde) implica em similaridade estrutural entre elas. Os métodos correntes de modelagem de proteínas por homologia implicam basicamente em quatro passos sucessivos: identificação e seleção de proteínas-molde; alinhamento das seqüências de resíduos; construção das coordenadas do modelo; validação.
 pelo método da homologia baseia-se no conceito de evolução molecular. Isto é, parte-se do princípio de que a semelhança entre as estruturas primárias desta proteína e de proteínas homólogas de estruturas tridimensionais conhecidas (proteínas-molde) implica em similaridade estrutural entre elas. Os métodos correntes de modelagem de proteínas por homologia implicam basicamente em quatro passos sucessivos: identificação e seleção de proteínas-molde; alinhamento das seqüências de resíduos; construção das coordenadas do modelo; validação.")
32
Threading Esta técnica é baseada na comparação da proteína em questão com modelos descritivos dos enovelamentos de proteínas homólogas Nesses modelos são descritas: a distância entre os resíduos de aminoácidos a estrutura secundária de cada fragmento as características fisico-químicas de cada resíduo

33
Ab Initio Entretanto, um grande desejo dos que trabalham com proteínas é o desenvolvimento de programas realmente eficientes para a modelagem ab initio Um programa que seja capaz de predizer a estrutura terciária de uma proteína, tendo como informação apenas a seqüência dos resíduos de aminoácidos e suas interações fisico-químicas, entre si e com o meio. Programas assim existem hoje mas têm muito a melhorar para que possamos confiar unicamente no seu resultado.

34
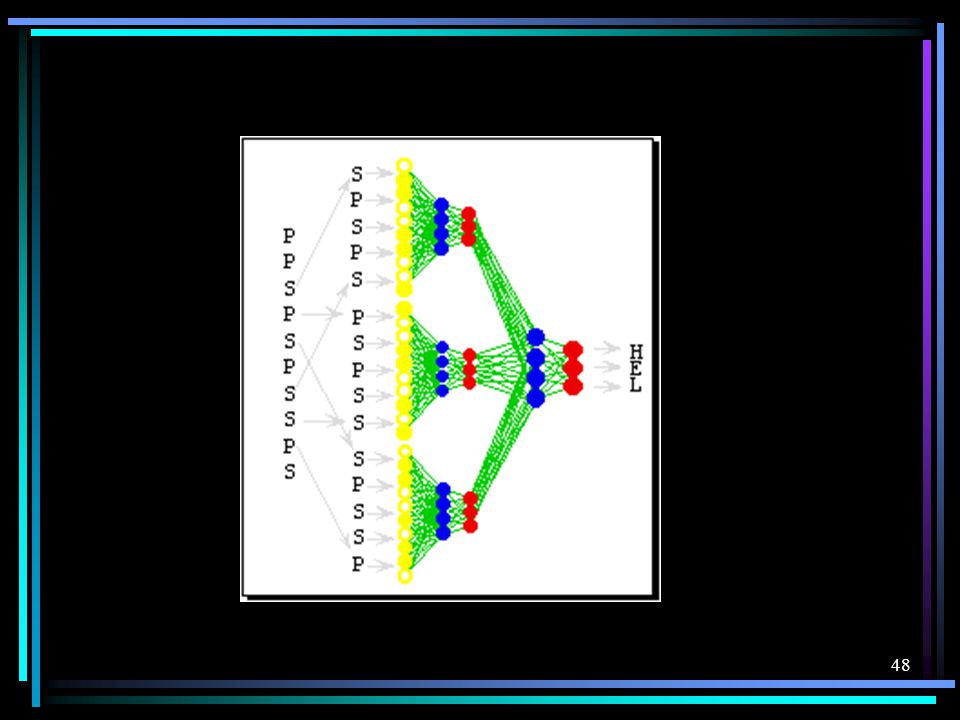
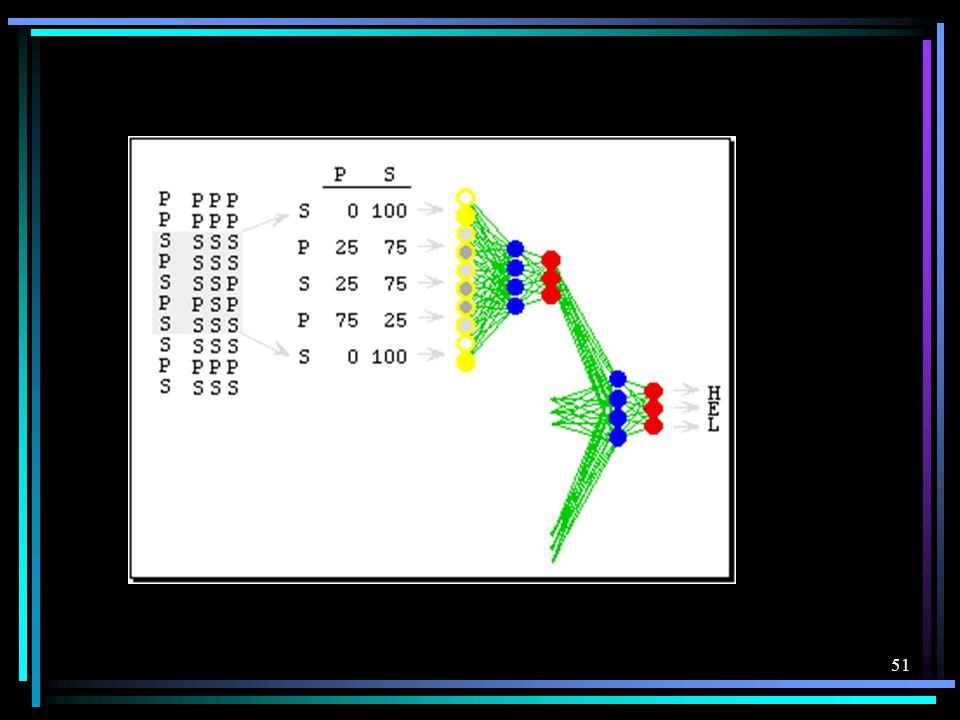
Predição de Estrutura Decomposição em três problemas:
Da Estrutura Primária para a Estrutura Secundária e outras Características Estruturais Da Estrutura Primária e Características Estruturais para Representações Topológicas De Representações Topológicas para Coordenadas 3D.

35
Protein Structure Terms
Protein Folds: The core 3D structure of a domain is called a fold. There are only a few thousand possible folds. Motif: A short conserved region in a protein sequence. Motifs are frequently highly conserved parts of domains. Domain: An independently folded unit within a protein, often joined by a flexible segment of the polypeptide chain. Class:used to classify protein domains according to their secondary structural content and organization Core:portion of the folded protein molecule that compromises the hydrophobic interior of the a helices and b sheets. Profile:a scoring matrix that represents a multiple sequence alignment of a protein family high-throughput protein structure determination: The goal of the current wave of structural genomics companies is to accelerate protein structure determination. Currently, only a few places can solve even several structures a year. No one has the technology in place to solve hundreds of crystal structures for hundreds of new proteins; rather, they will be solving folds. protein folding problem: Lies at the heart of a huge amount of modern biomedical research): the fact that thousands of different sequences can all form the same three- dimensional structure. protein taxonomy: The enormous variability in natural amino acid sequences does not correspond to a similar variability in conformations. The tertiary structure of protein show regularities, and many proteins have the same or similar folds, even in cases when they have no obvious evolutionary relation. Structure determination of proteins requires much more work than the determination of DNA sequences, but new structures appear at an increasing rate. With the growing database of known protein conformations there is a need to find ways of classifying protein, to define a taxonomy for proteins (or protein domains).
: the. fact that thousands of different sequences can all form the same three- dimensional structure. protein taxonomy: The enormous variability in natural amino acid sequences does not correspond. to a similar variability in conformations. The tertiary structure of protein show regularities, and many. proteins have the same or similar folds, even in cases when they have no obvious evolutionary. relation. Structure determination of proteins requires much more work than the determination of. DNA sequences, but new structures appear at an increasing rate. With the growing database of. known protein conformations there is a need to find ways of classifying protein, to define a. taxonomy for proteins (or protein domains).")
36
Protein Structure Terminology
a helix – the most abundant type of secondary structure in proteins. The helix has an average of 3.6 amino acids per turn with a hydrogen bond formed about every fourth residue. Average length is 10 amino acids b sheet- formed by hydrogen bonds between an average of 5-10 consecutive amino acids in one portion of the chain with another 5-10 further down the chain. The interacting regions may be adjacent, with a short loop in between or far apart with other structures in between.

37
Alpha Helix

39
Beta Sheet

41
Loops between alpha helix and beta sheets

42
Secondary Structure and Folding Classes
In the absence of “known” information about secondary structure, there are methods available for predicting the ability of a sequence to form a helices and b strands. Methods rely on observations made from groups of proteins whose three-dimensional structure has been experimentally determined Classification system based on the order of secondary structural elements within a protein

43
Secondary Structure Prediction
Predict the secondary structural conformation of each residue of protein sequences in general - making use of global rules applying across all sequence families (not those within individual families). Prediction programs are trained on data sets of non-homologous proteins of known structure (eg all sequence identity < 25%)
. Prediction programs are trained on data sets of non-homologous proteins of known structure (eg all sequence identity < 25%)")
44
Estruturas Secundárias
DSSP classes: H = alpha helix E = sheet G = 3-10 helix S = kind of turn T = beta turn B = beta bridge I = pi-helix (very rare) C = the rest CASP (harder) assignment: α = H and G β = E and B γ = the rest Alternative assignment: α = H β = B
 C = the rest. CASP (harder) assignment: α = H and G. β = E and B. γ = the rest. Alternative assignment: α = H. β = B.")
45
Algorithms: Nearest Neighbour - find the most similar sub-sequences of known structure (eg Levin, Robson, Garnier, 1986) Statistical, such as pairwise frequencies of amino acids as a function of separation and secondary structure (Garnier, Osguthorpe, Robson, 1978) Neural Networks, (eg PHD - Rost and Sander, 1993) Hybrid methods, eg using statistics, physico-chemical properties such as hydrophobic moments and others (eg DSC, King and Sternberg, 1996)
 Neural Networks, (eg PHD - Rost and Sander, 1993) Hybrid methods, eg using statistics, physico-chemical properties such as hydrophobic moments and others (eg DSC, King and Sternberg, 1996)")
46
History: The first generation prediction methods following in the 60's and 70's all based on single amino acid propensities The second-generation methods dominating the scene until the early 90's utilised propensities for segments of adjacent residues It seemed that prediction accuracy stalled at levels slightly above 60% The reason for this limit was the restriction to local information Can we introduce some global information into local stretches of residues

47
Traditional

49
Secondary structure prediction profits from divergence
Early on Dickerson [1976] realised that information contained in multiple alignments can improve predictions However, the breakthrough of the third generation methods to levels above 70% accuracy required a combination of larger databases with more advanced algorithms The major component of these new methods was the use of evolutionary information. All naturally evolved proteins with more than 35% pairwise identical residues over more than 100 aligned residues have similar structures

50
New database searches extend family divergence found
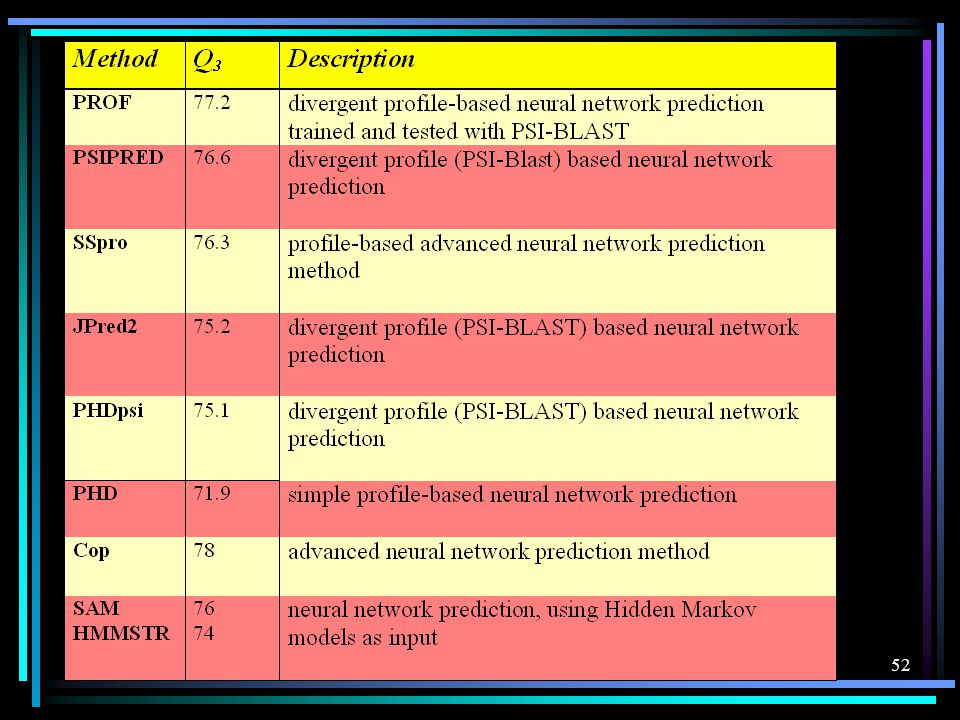
The breakthrough to large-scale routine searches has been achieved by the development of PSI-BLAST [Altschul, S. et al. (1997)] and Hidden Markov models [Eddy, S. R. (1998); Karplus, K., Barrett, C. & Hughey, R. (1998)] More data + refined search = better prediction Prediction accuracy peaks at 76% accuracy. The currently best methods reach a level of 76% three-state per-residue accuracy ( Table 1 ). This constitutes a sustained level more than four percentage points above last century's best method not using diverged profiles (PHD in Table 1 )
] and Hidden Markov models [Eddy, S. R. (1998); Karplus, K., Barrett, C. & Hughey, R. (1998)] More data + refined search = better prediction. Prediction accuracy peaks at 76% accuracy. The currently best methods reach a level of 76% three-state per-residue accuracy ( Table 1 ). This constitutes a sustained level more than four percentage points above last century s best method not using diverged profiles (PHD in Table 1 )")
53
Caution: over-optimism
Seemingly improve accuracy by ignoring short segments. There are many ways to publish higher levels of accuracy Comparing apples and oranges, or too few apples with one another There is NO value in comparing methods evaluated on different data sets For example, 16 new protein structures are clearly too few! For that set, JPred2, PHD, PROF, PSIPRED, SAM- T99sec and SSpro are indistinguishable Seemingly achieve 100% accuracy by using correlated sets EVA: automatic evaluation of automatic prediction servers

54
Clever methods can be more accurate 1/4
SSpro: advanced recursive neural network system The only method published recently that appears to improve prediction accuracy significantly not through more divergent profiles but through the particular algorithm is SSpro [Baldi, P., Brunak, S., Frasconi, P., Soda, G. & Pollastri, G. (1999)] The system never learns that secondary structure correlates between adjacent residues PHD addressed this problem by a second level structure-to-structure network that was trained on the predicted secondary structure from the first level sequence-to-structure network [Rost, B. & Sander, C. (1993)]. PSIPRED and JPred2 as well. Pierre Baldi and colleagues deviated substantially from this concept. Instead of using an additional network, they embedded the correlation into one single recursive neural network
] The system never learns that secondary structure correlates between adjacent residues. PHD addressed this problem by a second level structure-to-structure network that was trained on the predicted secondary structure from the first level sequence-to-structure network [Rost, B. & Sander, C. (1993)]. PSIPRED and JPred2 as well. Pierre Baldi and colleagues deviated substantially from this concept. Instead of using an additional network, they embedded the correlation into one single recursive neural network.")
55
Clever methods can be more accurate 2/4
HMMSTR: hidden Markov models for connecting library of structure fragments Can we predict secondary structure for protein U by local sequence similarity to segments of known structures {S} even when overall U differs from any of the known structures {S}? Yes, as shown by many nearest-neighbour-based prediction methods, the most successful of which seems to be NSSP [Salamov, A. A. & Solovyev, V. V. (1997)] A conceptually quite different realisation of the same concept has been implemented in HMMSTR by Chris Bystroff, David Baker and colleagues (2000)
] A conceptually quite different realisation of the same concept has been implemented in HMMSTR by Chris Bystroff, David Baker and colleagues (2000)")
56
Clever methods can be more accurate 3/4
HMMSTR: hidden Markov models for connecting library of structure fragments Firstly, build a library of local stretches (3-19) of residues with 'basic structural motifs' (I-sites) Secondly, assemble these local motifs through Hidden Markov models introducing structural context on the level of super-secondary structure Thus, the goal is to predict protein structure through identification of 'grammatical units of protein structure formation’ Although HMMSTR intrinsically aims at predicting higher order aspects of 3D structure, a side-result is the prediction of 1D secondary structure
 of residues with basic structural motifs (I-sites) Secondly, assemble these local motifs through Hidden Markov models introducing structural context on the level of super-secondary structure. Thus, the goal is to predict protein structure through identification of grammatical units of protein structure formation’ Although HMMSTR intrinsically aims at predicting higher order aspects of 3D structure, a side-result is the prediction of 1D secondary structure.")
57
Clever methods can be more accurate 4/4
Copenhagen: a Danish group developed a neural network-based method that is most amazing in many respects Petersen, T. N. et al. (2000). The authors estimate the method to yield levels above 77% prediction accuracy If true, this is the best current method Like PSIPRED, JPred2, and PROF, the method uses PSI-BLAST profiles as input, and like most methods since PHD a two-level approach addressing the problem of predicting short segments It replaces the standard 3 output units (for helix, strand, other), by 9 output units Also new is the particular way of weighting the average over different networks by the overall reliability of the prediction for that network, and the mere number of different networks considered (up to 800!)
. The authors estimate the method to yield levels above 77% prediction accuracy. If true, this is the best current method. Like PSIPRED, JPred2, and PROF, the method uses PSI-BLAST profiles as input, and like most methods since PHD a two-level approach addressing the problem of predicting short segments. It replaces the standard 3 output units (for helix, strand, other), by 9 output units. Also new is the particular way of weighting the average over different networks by the overall reliability of the prediction for that network, and the mere number of different networks considered (up to 800!)")
58
Combining mediocre and good methods is best
Combination improves on non-systematic errors Systematic errors, e.g., through non-local effects White noise errors caused by, e.g., the succession of the examples during training neural networks Theoretically, combining any number of methods improves accuracy as long as the errors of the individual methods are mutually independent and are not only systematic PHD - and more recently others [Chandonia, JPred2, Copenhagen] - utilised this fact by combining different neural networks.

59
Discussion Methods improved significantly over last two years
Growing databases and improved search techniques - predominantly through the iterated PSI-BLAST tool - yielded a substantial improvement in secondary structure prediction accuracy over the last two years. State-of-the-art methods now reach sustained levels of 76% prediction accuracy What is the limit of prediction accuracy? 88% is the limit, but shall we ever reach close to there? Larger databases may get us six percentage points higher, and it may not. The answer remains nebulous

60
References B. Rost (2001) Protein secondary structure prediction continues to rise. Journal of Structural Biology, 134, pp (Columbia University). Bystroff, C., Thorsson, V. & Baker, D. (2000). HMMSTR: a hidden Markov model for local sequence-structure correlations in proteins. J. Mol. Biol., 301, (University of Washington) Cuff, J. A., Clamp, M. E., Siddiqui, A. S., Finlay, M. & Barton, G. J. (1998). JPred: a consensus secondary structure prediction server. Bioinformatics, 14, (JPred – Oxford/Cambridge) Cuff, J. A. & Barton, G. J. (2000). Application of multiple sequence alignment profiles to improve protein secondary structure prediction. Proteins, 40, (JPred2) Rost, B. (1996). PHD: predicting one-dimensional protein structure by profile based neural networks. Meth. Enzymol., 266, (PHD – Heidelberg – Germany)
 Protein secondary structure prediction continues to rise. Journal of Structural Biology, 134, pp (Columbia University). Bystroff, C., Thorsson, V. & Baker, D. (2000). HMMSTR: a hidden Markov model for local sequence-structure correlations in proteins. J. Mol. Biol., 301, (University of Washington) Cuff, J. A., Clamp, M. E., Siddiqui, A. S., Finlay, M. & Barton, G. J. (1998). JPred: a consensus secondary structure prediction server. Bioinformatics, 14, (JPred – Oxford/Cambridge) Cuff, J. A. & Barton, G. J. (2000). Application of multiple sequence alignment profiles to improve protein secondary structure prediction. Proteins, 40, (JPred2) Rost, B. (1996). PHD: predicting one-dimensional protein structure by profile based neural networks. Meth. Enzymol., 266, (PHD – Heidelberg – Germany)")
61
References Przybylski, D. & Rost, B. (2000). PSI-BLAST for structure prediction: plug-in and win. Columbia University (PHDPsi) Rost WWW, B. (2000). Better secondary structure prediction through more data. Columbia University, WWW document ( (PROF) Altschul, S., Madden, T., Shaffer, A., Zhang, J., Zhang, Z. et al. (1997). Gapped Blast and PSI-Blast: a new generation of protein database search programs. Nucl. Acids Res., 25, (PSI-BLAST – USA) Jones, D. T. (1999). Protein secondary structure prediction based on position-specific scoring matrices. J. Mol. Biol., 292, (PSIPRED – Warwick) Karplus, K., Barrett, C. & Hughey, R. (1998). Hidden Markov models for detecting remote protein homologies. Bioinformatics, 14, (SAM-T99Sec – University of California Sta. Cruz)
. Better secondary structure prediction through more data. Columbia University, WWW document ( (PROF) Altschul, S., Madden, T., Shaffer, A., Zhang, J., Zhang, Z. et al. (1997). Gapped Blast and PSI-Blast: a new generation of protein database search programs. Nucl. Acids Res., 25, (PSI-BLAST – USA) Jones, D. T. (1999). Protein secondary structure prediction based on position-specific scoring matrices. J. Mol. Biol., 292, (PSIPRED – Warwick) Karplus, K., Barrett, C. & Hughey, R. (1998). Hidden Markov models for detecting remote protein homologies. Bioinformatics, 14, (SAM-T99Sec – University of California Sta. Cruz)")
62
References Baldi, P., Brunak, S., Frasconi, P., Soda, G. & Pollastri, G. (1999). Exploiting the past and the future in protein secondary structure prediction. Bioinformatics, 15, (Sspro – University of California at Irvine and Italy) Petersen, T. N., Lundegaard, C., Nielsen, M., Bohr, H., Bohr, J. et al. (2000). Prediction of protein secondary structure at 80% accuracy. Proteins, 41, – Denamark, including Brunak) Salamov, A. A. & Solovyev, V. V. (1997). Protein secondary structure prediction using local alignments. J. Mol. Biol., 268, (nearest-neigbour method)
. Exploiting the past and the future in protein secondary structure prediction. Bioinformatics, 15, (Sspro – University of California at Irvine and Italy) Petersen, T. N., Lundegaard, C., Nielsen, M., Bohr, H., Bohr, J. et al. (2000). Prediction of protein secondary structure at 80% accuracy. Proteins, 41, – Denamark, including Brunak) Salamov, A. A. & Solovyev, V. V. (1997). Protein secondary structure prediction using local alignments. J. Mol. Biol., 268, (nearest-neigbour method)")
Apresentações semelhantes





. Nenhuns direitos reservados, excepto para fins comerciais. Por favor, não coloque.>")













