Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Atualidades em Cólera Aviária
Thales Quedi Furian 5 de Novembro Simpósio de Sanidade Avícola (AVISULAT/UFSM) IV Congresso Sul Brasileiro de Avicultura, Suinocultura e Laticínios
 IV Congresso Sul Brasileiro de Avicultura, Suinocultura e Laticínios.")
2
Cólera Aviária Uma das patologias aviárias mais antigas;
Diagnóstico diferencial de enfermidades com notificação obrigatória; Casos frequentes entre pequenas criações independentes

3
Cólera Aviária Mas também é uma preocupação para a avicultura industrial.... Grande concentração de unidades de produção Alta densidade populacional aviários + (Fotos: SOARES, 2009; JÚNIOR, 2009)
")
4
Cólera Aviária Baixos níveis de biosseguridade
Mas também é uma preocupação para a avicultura industrial.... Grande concentração de unidades de produção Alta densidade populacional aviários Baixos níveis de biosseguridade (Fotos: SOARES, 2009; JÚNIOR, 2009)
")
5
Cólera Aviária Baixos níveis de biosseguridade
Mas também é uma preocupação para a avicultura industrial.... Grande concentração de unidades de produção Alta densidade populacional aviários Doenças respiratórias Baixos níveis de biosseguridade (Fotos: SOARES, 2009; JÚNIOR, 2009)
")
6
Cólera Aviária Baixos níveis de biosseguridade
Diferentes hospedeiros e cronicamente infectados Mas também é uma preocupação para a avicultura industrial.... Grande concentração de unidades de produção Alta densidade populacional aviários Doenças respiratórias Baixos níveis de biosseguridade (Fotos: SOARES, 2009; JÚNIOR, 2009)
")
7
Família Pasteurellaceae
(HARPER et al., 2006) Denominação do gênero em homenagem a Louis Pasteur 10 gêneros (Pasteurella, Actinobacillus, Haemophilus, Avibacterium); (MUHAIRWWA et al., 2001; SNIPES et al., 2002; LEOTA et al., 2006, NASCIMENTO et al., 2009) - constantes reclassificações taxonômicas; “A bactéria Pasteurella multocida é um patógeno enigmático” (WILKIE et al., 2012) número de síndromes relacionadas e de hospedeiros acometidos; agente primário (rinite atrófica, septicemia hemorrágica, cólera aviária)
 Denominação do gênero em. homenagem a Louis Pasteur. 10 gêneros (Pasteurella, Actinobacillus, Haemophilus, Avibacterium); (MUHAIRWWA et al., 2001; SNIPES et al., 2002; LEOTA et al., 2006, NASCIMENTO et al., 2009) - constantes reclassificações taxonômicas; A bactéria Pasteurella multocida é um patógeno enigmático (WILKIE et al., 2012) número de síndromes relacionadas e de hospedeiros acometidos; agente primário (rinite atrófica, septicemia hemorrágica, cólera aviária)")
8
Pasteurella multocida
Figura 1. Micrografia eletrônica – Pasteurella spp Figura 2. Coloração esfregaço sanguineo – P. multocida Bacilo Gram negativo; Imóvel, não formador de esporos; Coloração bipolar típica; Anaeróbio facultativo; (CHRISTENSEN; BISGAARD, 2006; GLISSON, 2008 )
")
9
Cólera Aviária Forma aguda Forma crônica
anorexia, cianose, estertores, descargas nasais, diarréia, morte súbita; petéquias, hemorragias, ausência lesões; Cólera Aviária lesões edematosas/inflamatórias associadas com o local de infecção; lesões no sistema reprodutivo; Forma aguda Forma crônica Fotos: CORNELL UNIVERSITY, CDPA/UFRGS)
")
10
Epidemiologia Aves entre 16-40 semanas mais susceptíveis
Lotes clinicamente recuperados Reservatório do agente Aves aquáticas Mamíferos (PETERSEN et al., 2001) (SAMUEL et al., 2007) (MUHAURWA et al., 2001) (WILKIE et al., 2012) Aves silvestres Descrita em mais de 100 espécies
 (SAMUEL et al., 2007) (MUHAURWA et al., 2001) (WILKIE et al., 2012) Aves silvestres. Descrita em mais de 100 espécies.")
11
Como realizar o diagnóstico laboratorial atualmente?

12
Isolamento convencional
BHI Ágar Sangue 5% sangue ovino desf. Ágar MacConkey 37o C 24 horas Testes bioquímicos Teste Catalase Teste Oxidase Colônias compatíveis anaerobiose aerobiose Coloração Gram/ Azul de metileno Fígado Pulmão Sangue cardíaco Medula óssea (A laboratory manual for the isolation, identification and characterization of avian pathogens - GLISSON et al., 2008)
")
13
Diagnóstico Meios de isolamento: ágar sangue, TSA, DSA
(Fonte: CDPA/UFRGS, 2008) Figura 3. Colônias de Pasteurella multocida isoladas de diferentes hospedeiros Aves - UFRGS Bovinos - UFRGS Suínos - UFRGS ATCC 29545 Meios de isolamento: ágar sangue, TSA, DSA - 5% soro sanguíneo; Colônias geralmente acinzentadas e lisas; Inoculação em camundongos; - morte em 24 a 48 horas; (CHRISTENSEN et al., 2007)
 Figura 3. Colônias de Pasteurella multocida isoladas de diferentes hospedeiros. Aves - UFRGS. Bovinos - UFRGS. Suínos - UFRGS. ATCC Meios de isolamento: ágar sangue, TSA, DSA. - 5% soro sanguíneo; Colônias geralmente acinzentadas e lisas; Inoculação em camundongos; - morte em 24 a 48 horas; (CHRISTENSEN et al., 2007)")
14
Diagnóstico Algumas dificuldades entre os testes fenotípicos
- Isolamento regiões contaminadas; - Período para obtenção de subcultivos puros; - Questão bem estar animal; Métodos moleculares de diagnóstico Detecção do gene pls kmt (KASTEN et al., 1997) (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes pm0762/pm231 Ensaio 5’ Taq nuclease (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método extração DNA
 (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes. pm0762/pm231. Ensaio. 5’ Taq nuclease. (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método. extração DNA.")
15
Diagnóstico Algumas dificuldades entre os testes fenotípicos
- Isolamento regiões contaminadas; - Período para obtenção de subcultivos puros; - Questão bem estar animal; Métodos moleculares de diagnóstico Detecção do gene pls kmt (KASTEN et al., 1997) (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes pm0762/pm231 Ensaio 5’ Taq nuclease (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método extração DNA
 (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes. pm0762/pm231. Ensaio. 5’ Taq nuclease. (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método. extração DNA.")
16
Diagnóstico Algumas dificuldades entre os testes fenotípicos
- Isolamento regiões contaminadas; - Período para obtenção de subcultivos puros; - Questão bem estar animal; Métodos moleculares de diagnóstico Detecção do gene pls kmt (KASTEN et al., 1997) (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes pm0762/pm231 Ensaio 5’ Taq nuclease (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método extração DNA Preservação Amostras liofilizadas
 (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes. pm0762/pm231. Ensaio. 5’ Taq nuclease. (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método. extração DNA. Preservação. Amostras liofilizadas.")
17
Diagnóstico Algumas dificuldades entre os testes fenotípicos
- Isolamento regiões contaminadas; - Período para obtenção de subcultivos puros; - Questão bem estar animal; Métodos moleculares de diagnóstico Detecção do gene pls kmt (KASTEN et al., 1997) (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes pm0762/pm231 Ensaio 5’ Taq nuclease (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método extração DNA Preservação Congeladas a -80oC sangue total ovino
 (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes. pm0762/pm231. Ensaio. 5’ Taq nuclease. (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método. extração DNA. Preservação. Congeladas a -80oC. sangue total ovino.")
18
Diagnóstico Algumas dificuldades entre os testes fenotípicos
- Isolamento regiões contaminadas; - Período para obtenção de subcultivos puros; - Questão bem estar animal; Métodos moleculares de diagnóstico Detecção do gene pls kmt (KASTEN et al., 1997) (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes pm0762/pm231 Ensaio 5’ Taq nuclease (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método extração DNA Preservação Congeladas a -80oC soro bovino + 7,5% de glicose (LAPAGE & REDWAY, 1974)
 (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes. pm0762/pm231. Ensaio. 5’ Taq nuclease. (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método. extração DNA. Preservação. Congeladas a -80oC. soro bovino + 7,5% de glicose. (LAPAGE & REDWAY, 1974)")
19
Diagnóstico Algumas dificuldades entre os testes fenotípicos
- Isolamento regiões contaminadas; - Período para obtenção de subcultivos puros; - Questão bem estar animal; Métodos moleculares de diagnóstico Detecção do gene pls kmt (KASTEN et al., 1997) (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes pm0762/pm231 Ensaio 5’ Taq nuclease (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método extração DNA Preservação Congeladas a -80oC BHI+ glicerol
 (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes. pm0762/pm231. Ensaio. 5’ Taq nuclease. (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método. extração DNA. Preservação. Congeladas a -80oC. BHI+ glicerol.")
20
Diagnóstico Algumas dificuldades entre os testes fenotípicos
- Isolamento regiões contaminadas; - Período para obtenção de subcultivos puros; - Questão bem estar animal; Métodos moleculares de diagnóstico Detecção do gene pls kmt (KASTEN et al., 1997) (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes pm0762/pm231 Ensaio 5’ Taq nuclease (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método extração DNA Preservação Congeladas a -20oC BHI+ glicerol soro bovino + 7,5% de glicose sangue total ovino
 (LEE et al., 2000) (LIU et al., 2004) Detecção dos genes. pm0762/pm231. Ensaio. 5’ Taq nuclease. (CORNEY et al., 2007) (TOWNSEND et al., 1998) Variação método. extração DNA. Preservação. Congeladas a -20oC. BHI+ glicerol. soro bovino + 7,5% de glicose. sangue total ovino.")
21
Cólera Aviária Patogenia Habitante normal das vias aéreas
Agente Ambiente Hospedeiro Manejo inadequado Nutrição deficiente Infestação parasitária Infecções concomitantes (MUHAIRWA et al., 2000; SAMUEL et al., 2007; WILKIE et al., 2012) Trato respiratório superior Principal sítio Infecção do pulmão e sacos aéreos Disseminação para diferentes tecidos Corrente sanguínea Morte Choque septicêmico
 Trato respiratório superior. Principal sítio. Infecção do pulmão. e sacos aéreos. Disseminação para. diferentes tecidos. Corrente sanguínea. Morte. Choque septicêmico.")
22
? Após mais de 125 anos dos trabalhos desenvolvidos por Pasteur...
Patogenia e a virulência das cepas Patogenia e a virulência das cepas Sítio inicial de colonização Vacinação ideal Patogenicidade das cepas ? Invasão da mucosas Vacinas vivas e inativadas Microrganismos oportunistas em vertebrados (CHRISTENSEN; BISGAARD, 2006; DZIVA et al., 2008; HARPER et al., 2010)
")
24
Acesso em 10/10 3511 document results

26
Acesso em 10/10 1167 document results

28
Acesso em 10/10 406 document results

29
“A manipulação genética de P
“A manipulação genética de P. multocida deve auxiliar na elucidação dos mecanismos patogênicos e da associação com diferentes hospedeiros.” Ian Wilkie – 2012 Department of Microbiology Australian Research Council Centre of Excellence in Structural and Functional Microbial Genomics, Monash University, Australia

30
primeiro sequenciamento completo do genoma em 2001;
menor uso de ferramentas de manipulação/análise genética nos últimos 25 anos; primeiro sequenciamento completo do genoma em 2001; (May et al., 2001) Tabela 1. Característica do genoma das cepas de Pasteurella multocida sequenciadas Cepa Completo Hospedeiro/ tecido Genoma (Mbp) G+C Pm70 sim ave 2.26 40.40 36950 bovino/pulmão 2.35 40.44 Anand1G não caprino 2.29 40.52 Anand1P ave/fígado 2.02 40.22 P3480 suíno/pulmão 2.08 40.15 X73 2.31 40.31 VP161 40.21 P903 suíno 40.14 M1404 Bovino 2.30 40.27 Isolados de CA Boyce et al., 2012
 Tabela 1. Característica do genoma das cepas de Pasteurella multocida sequenciadas. Cepa. Completo. Hospedeiro/ tecido. Genoma (Mbp) G+C. Pm70. sim. ave bovino/pulmão Anand1G. não. caprino Anand1P. ave/fígado P3480. suíno/pulmão X VP P903. suíno M1404. Bovino Isolados de CA. Boyce et al.,")
31
Análise filogenética Todas as cepas são fortemente relacionadas;
Não há correlação entre o parentesco filogenético e o local de isolamento, o sorogrupo ou o hospedeiro de origem das amostras;

32
Como e por que classificar os isolados de P. multocida?

33
Testes fenotípicos para classificação da subespécie e do biovar
Biotipificação: fator chave na identificação de P. multocida; P. multocida subsp. multocida subsp. septica subsp. gallicida (DZIVA et al., 2008 ) Tabela 2. Características bioquímicas para diferenciação de subespécies de Pasteurella multocida Teste P. multocida P. septica P. gallicida Dulcitol - + Sorbitol (MUTTERS et al., 1985 ) Enzima α-glicosidase e sequenciamento de genes housekeeping; (HUNT et al., 2001; KUHNERT, P & KORCZAK, B.M., 2006 )
 Tabela 2. Características bioquímicas para diferenciação de subespécies de Pasteurella multocida. Teste. P. multocida. P. septica. P. gallicida. Dulcitol. - + Sorbitol. (MUTTERS et al., 1985 ) Enzima α-glicosidase e sequenciamento de genes housekeeping; (HUNT et al., 2001; KUHNERT, P & KORCZAK, B.M., 2006 )")
34
Testes fenotípicos para classificação da subespécie e do biovar
Classificação das amostras em 13 diferentes biovares Tabela 3. Características bioquímicas dos biovares de Pasteurella multocida Fermentação 1 2 3 4 5 6 7 8 9 10 12 13 14 Xilose - + Maltose Ornitina Trehalose Sorbitol Dulcitol Lactose (FEGAN et al., 1995; BLACKALL et al., 1997)
")
35
Levantamento – região sul do Brasil isolados de cólera aviária
Subespécie Percentual (%) multocida 87,50 septica 10,71 gallicida 1,79 Total 100 Biovar Percentual (%) 1 8,93 2 7,14 3 35,71 4 1,79 5 3,57 7 8 9 12,50 10 13 23,21 NT 0,00 Total 100 (Dados não publicados, 2014)
 multocida. 87,50. septica. 10,71. gallicida. 1,79. Total Biovar. Percentual (%) 1. 8, , , , , , ,21. NT. 0,00. Total (Dados não publicados, 2014)")
36
Levantamento – região sul do Brasil isolados de cólera aviária
Subespécie Percentual (%) multocida 87,50 septica 10,71 gallicida 1,79 Total 100 Biovar Percentual (%) 1 8,93 2 7,14 3 35,71 4 1,79 5 3,57 7 8 9 12,50 10 13 23,21 NT 0,00 Total 100 MUHAIRWA et al., 2001 PEDERSEN et al., 2003 LEOTTA et al., 2006 (Dados não publicados, 2014)
 multocida. 87,50. septica. 10,71. gallicida. 1,79. Total Biovar. Percentual (%) 1. 8, , , , , , ,21. NT. 0,00. Total MUHAIRWA et al., PEDERSEN et al., LEOTTA et al., (Dados não publicados, 2014)")
37
Levantamento – região sul do Brasil isolados de cólera aviária
Subespécie Percentual (%) multocida 87,50 septica 10,71 gallicida 1,79 Total 100 Biovar Percentual (%) 1 8,93 2 7,14 3 35,71 4 1,79 5 3,57 7 8 9 12,50 10 13 23,21 NT 0,00 Total 100 FEGAN et al., 1995 VARGA et al., 2013 (Dados não publicados, 2014)
 multocida. 87,50. septica. 10,71. gallicida. 1,79. Total Biovar. Percentual (%) 1. 8, , , , , , ,21. NT. 0,00. Total FEGAN et al., VARGA et al., (Dados não publicados, 2014)")
38
Biovar 2: hgbB-; pfhA+ Biovar 3: hgbB+; pfhA- Biovar Percentual (%) 1 8,93 2 7,14 3 35,71 4 1,79 5 3,57 7 8 9 12,50 10 13 23,21 NT 0,00 Total 100 Não houve associação entre os biovares e a presença de 23 genes de virulência pesquisados (p˃ 0,05) (Dados não publicados, 2014)
 (Dados não publicados, 2014)")
39
Testes fenotípicos para classificação da subespécie e do biovar
Limitações Grande número de isolados não tipificáveis; Variações de reprodutibilidade dos testes; Menor poder discriminatório Combinada com técnicas moleculares de tipificação (MATSUMOTO & STRAIN; 1993; VARGA et al., 2007; DZIVA et al., 2008 ; GARCIA et al., 2011)
")
40
Tipificação molecular
Testes moleculares X testes de biotipificação Tipificação molecular Tipificação: isolados compartilham propriedades que os diferenciam;

41
Tipificação molecular
Testes moleculares X testes de biotipificação Análise de enzima de restrição - REA Ribotipagem Análise do Polimorfismo de DNA Amplificado ao Acaso - RAPD Sequência Palindrômica Repetitiva - REP Análise do Polimorfismo no comprimento de fragmentos de restrição - RFLP Tipificação por sequenciamento de múltiplos loci - MLST Diferenciação de cepas de P. multocida ( MUHAIRWA et al., 2001; CHRISTENSEN & BISGAARD, 2006)
")
42
Tipificação molecular
Técnica de PCR-RFLP em P. multocida; - discriminação do gene ompH; - estrutura do lipopolissacarídeo; (TSAI et al., 2011) (BOROWSKI et al., 2001; JABBARI et al., 2005; SELLYEI et al., 2013) Levantamento – região sul do Brasil isolados de cólera aviária Amplificação dos genes ompH (Sellyei et al., 2013); oma87 (Furian et al., 2013);
 (BOROWSKI et al., 2001; JABBARI et al., 2005; SELLYEI et al., 2013) Levantamento – região sul do Brasil isolados de cólera aviária. Amplificação dos genes ompH (Sellyei et al., 2013); oma87 (Furian et al., 2013);")
43
I VI V VII II PM GenBank U50907 – ompH gene strain X-73
Figura 1. Eletroforese em gel de agarose 1% corado com brometo de etídeo - produto de amplificação gene ompH (aprox pb). A seta preenchida indica fragmento de 500pb. Restriction mapper version 3 DraI HindIIIPvuII 5’-TTT AAA-3’ 3’-AAA TTT-5’ 5’-A AGCT T-3’ 3’-T TCGA A -5’ 5’-GTPy PuAC-3’ 3’-CApu PyTG-5’ I VI V VII II PM Figura 2. Perfis gerados (I-VII) das cepas de Pasteurella multocida de origem aviária a partir da clivagem do produto de amplificação do gene ompH com as enzimas DraI e HindIII
. A seta preenchida indica fragmento de 500pb. Restriction mapper. version 3. DraI. HindIIIPvuII. 5’-TTT AAA-3’ 3’-AAA TTT-5’ 5’-A AGCT T-3’ 3’-T TCGA A -5’ 5’-GTPy PuAC-3’ 3’-CApu PyTG-5’ I. VI. V. VII. II. PM. Figura 2. Perfis gerados (I-VII) das cepas de Pasteurella multocida de origem aviária a partir da clivagem do produto de amplificação do gene ompH com as enzimas DraI e HindIII.")
44
PCR-RFLP Grupo genético Frequência relativa (%) I 12,28 II 42,11 III
Alta homologia de alguns perfis com determinados sorotipos; PCR-RFLP Associação do perfil VII com o biovar 13; Perfil II Sorotipo 3 Perfil VII Sorotipo 11 Sorotipo 3,4 Sorotipo 2 Perfil III Sorotipo 4 Sorotipo 9 Sorotipo 15 Sorotipo 5,16 Perfil V Sorotipo 10 Sorotipo 12 Sorotipo 14 Sorotipo 6 Sorotipo 7 Sorotipo 8 Sorotipo 13 Perfil VI Perfis gerados (I-VII) das cepas de Pasteurella multocida de origem aviária a partir da clivagem do produto de amplificação do gene ompH com as enzimas DraI e HindIII Grupo genético Frequência relativa (%) I 12,28 II 42,11 III 3,51 IV V 17,54 VI 8,77 VII Figura 3. Relação filogenética da sequência nucleotídica de 638pb do gene ompH entre os perfis gerados através de PCR-RFLP e os diferentes sorotipos de Pasteurella multocida
 das cepas de Pasteurella multocida de origem aviária a partir da clivagem do produto de amplificação do gene ompH com as enzimas DraI e HindIII. Grupo genético. Frequência. relativa (%) I. 12,28. II. 42,11. III. 3,51. IV. V. 17,54. VI. 8,77. VII. Figura 3. Relação filogenética da sequência nucleotídica de 638pb do gene ompH entre os perfis gerados através de PCR-RFLP e os diferentes sorotipos de Pasteurella multocida.")
45
Kruskall-Wallis test (p<0,01)
Distribuição do índice de patogenicidade versus perfil genético de isolados de origem aviária de Pasteurella multocida 6,05 8,97 5,81 4,03 8,77 5,62 Grupo genético IP médio I 6,05a II 8,97b III/IV 8,77b V 5,81a VI 4,03a VII 5,62a

46
Métodos de classificação de P
Métodos de classificação de P. multocida são empregados em estudos epidemiológicos e na diferenciação de cepas isoladas em surtos (DZIVA et al., 2008). A variabilidade antigênica dificulta o estabelecimento de uma vacina eficiente (DAVIES et al., 2003). Importante determinar os sorotipos de P. multocida: bacterinas protegem pobremente contra desafios das cepas heterólogas.
. A variabilidade antigênica dificulta o estabelecimento de uma vacina eficiente (DAVIES et al., 2003). Importante determinar os sorotipos de P. multocida: bacterinas protegem pobremente contra desafios das cepas heterólogas.")
47
(BISGAARD, 2008; HARPER et al., 2012)
Cápsula Cápsula e LPS: base para a classificação de P. multocida Classificadas em cinco sorogrupos (A, B, D, E, F) Classificadas em 16 sorovares Casos de CA predominantes: sorogrupo A e sorovares 1-3 ou 3-4; antígenos capsulares; antígenos somáticos; (BISGAARD, 2008; HARPER et al., 2012)
 Classificadas em 16 sorovares. Casos de CA predominantes: sorogrupo A e sorovares 1-3 ou 3-4; antígenos capsulares; antígenos somáticos; (BISGAARD, 2008; HARPER et al., 2012)")
48
Glicosaminoglicanos idênticos aos componentes da matriz celular
N-acetil-glicosamina Ácido glicurônico Ácido hialurônico Condroitina Glicosaminoglicanos idênticos aos componentes da matriz celular da célula eucariótica Pode não ocorrer resposta imune do hospedeiro Heparina (DAVIES et al., 2004; BOYCE et al., 2010)
")
49
Cápsula Importante papel na interação com o organismo infectado
cepas com cápsula X mutantes acapsulares Resistência à dessecação; Maior adesão (?); Atividade antifagocitária; Interação com o sistema complemento; (JACQUES et al., 1993; PRUIMBOOM et al., 1996; CHUNG et al., 2001,;BOYCE et al., 2010; HARPER et al., 2012)
; Atividade antifagocitária; Interação com o sistema complemento; (JACQUES et al., 1993; PRUIMBOOM et al., 1996; CHUNG et al., 2001,;BOYCE et al., 2010; HARPER et al., 2012)")
50
Métodos de tipificação capsular e relação com a patogenicidade
Classificação fundamental: - relação entre tipo capsular, patogenia e predisposição do hospedeiro a um sorogrupo; rinite atrófica septicemia hemorrágica cólera aviária tipo D tipo B ou E tipo A Prevalência pode variar conforme a distribuição geográfica e ao longo do tempo; (JAGLIC et al., 2005; WOO;KIM , 2006; HAPER et al., 2012)
")
51
Métodos de tipificação capsular e relação com a patogenicidade
Tipificação: teste de hemaglutinação indireta - isolados podem não aglutinar com antissoros homólogos; - depende da presença do antígeno capsular; Métodos fenotípicos não-sorológicos - Teste da hialuronidase: amostras do tipo A despolimerização do ácido hialurônico - Teste da acriflavina: amostras do tipo D floculação em caldo - microscopia com luz oblíqua colônias iridescentes - microscopia eletrônica espessura da cápsula (CARTER, 1955) patogenicidade (CARTER & RUNDELL, 1973; CARTER & SUBRONTO, 1973; GLISSON et al., 2008; BORRATHYBAY et al., 2003)
 patogenicidade. (CARTER & RUNDELL, 1973; CARTER & SUBRONTO, 1973; GLISSON et al., 2008; BORRATHYBAY et al., 2003)")
52
Tabela 1. Característica do genoma das cepas de Pasteurella multocida sequenciadas
Completo Hospedeiro/ tecido Genoma (Mbp) G+C Pm70 sim ave 2.26 40.40 36950 bovino/pulmão 2.35 40.44 Anand1G não caprino 2.29 40.52 Anand1P ave/fígado 2.02 40.22 P3480 suíno/pulmão 2.08 40.15 X73 2.31 40.31 VP161 40.21 P903 suíno 40.14 M1404 Bovino 2.30 40.27 locus de biossíntese da cápsula A:1 (Chung et al., 1998) locus de biossíntese da cápsula B:2 (Boyce et al., 2000) Boyce et al., 2012
 G+C. Pm70. sim. ave bovino/pulmão Anand1G. não. caprino Anand1P. ave/fígado P3480. suíno/pulmão X VP P903. suíno M1404. Bovino locus de biossíntese da cápsula A:1. (Chung et al., 1998) locus de biossíntese da cápsula B:2. (Boyce et al., 2000) Boyce et al.,")
54
Cápsula Região 01 Região 02 Região 03
Região 1: proteínas para transporte dos polissacarídeos da cápsula; Região 3: enzimas para fosforilação; Região 2: enzima para biossíntese dos monômeros de açúcar; (CHUNG et al., 1998, BOYCE et al., 2010) Região 01 Região 02 Região 03 hexA hexB hexC hexD hyaA hyaB hyaC hyaD hyaE phyA phyB Pasteurella multocida A:1 cexA cexB cexC cexD lipA bcbI bcbH bcbF lipB bcbG bcbE bcbD bcbC bcbB bcbA Pasteurella multocida B:2 (adapatado de: BOYCE et al., 2000)
 Região 01. Região 02. Região 03. hexA. hexB. hexC. hexD. hyaA. hyaB. hyaC. hyaD. hyaE. phyA. phyB. Pasteurella multocida A:1. cexA. cexB. cexC. cexD. lipA. bcbI. bcbH. bcbF. lipB. bcbG. bcbE. bcbD. bcbC. bcbB. bcbA. Pasteurella multocida B:2. (adapatado de: BOYCE et al., 2000)")
55
Tabela 4. Tipificação capsular de 54 isolados de Pasteurella multocida de origem aviária com testes fenotípicos não sorológicos e multiplex-PCR Análise Sorogrupo A (%) D (%) NT (%) Total (%) Testes fenotípicos 41 (75,93)a 2 (3,7)a 11 (20,37)a 54 (100) Multiplex-PCR 49 (90,74)b 3 (5,56)a 2 (3,7)b X2 (1) = 4,267 p= 0,034 (p<0,05) Leotta et al. (2006), Shivachandra et al. (2006), Jabbari et al. (2006)
 D (%) NT (%) Total (%) Testes fenotípicos. 41 (75,93)a. 2 (3,7)a. 11 (20,37)a. 54 (100) Multiplex-PCR. 49 (90,74)b. 3 (5,56)a. 2 (3,7)b. X2 (1) = 4,267 p= 0,034 (p<0,05) Leotta et al. (2006), Shivachandra et al. (2006), Jabbari et al. (2006)")
56
Fatores de virulência Cápsula e LPS Fímbrias e adesinas
Proteínas externas de membrana Toxina dermonecrótica (ptfA, pfhA, hsf-1, tadA) (ompH, oma87, ompA, plpB) (toxA) (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur); Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro;
 (ompH, oma87, ompA, plpB) (toxA) (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur); Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro;")
57
Fatores de virulência Cápsula e LPS Fímbrias e adesinas
FHA PtfA (adaptado de HATFALUDI et al., 2010) Fatores de virulência Cápsula e LPS Fímbrias e adesinas Proteínas externas de membrana Toxina dermonecrótica (ptfA, pfhA, hsf-1, tadA) (ompH, oma87, ompA, plpB) (toxA) (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur); Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro;
 Fatores de virulência. Cápsula e LPS. Fímbrias e adesinas. Proteínas externas de membrana. Toxina dermonecrótica. (ptfA, pfhA, hsf-1, tadA) (ompH, oma87, ompA, plpB) (toxA) (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur); Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro;")
58
Fatores de virulência Cápsula e LPS Fímbrias e adesinas
OmpH Oma87 (adaptado de HATFALUDI et al., 2010) Fatores de virulência Cápsula e LPS Fímbrias e adesinas Proteínas externas de membrana Toxina dermonecrótica (ptfA, pfhA, hsf-1, tadA) (ompH, oma87, ompA, plpB) (toxA) (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur); Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro;
 Fatores de virulência. Cápsula e LPS. Fímbrias e adesinas. Proteínas externas de membrana. Toxina dermonecrótica. (ptfA, pfhA, hsf-1, tadA) (ompH, oma87, ompA, plpB) (toxA) (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur); Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro;")
59
Fatores de virulência Cápsula e LPS Fímbrias e adesinas
Proteínas externas de membrana Toxina dermonecrótica (ptfA, pfhA, hsf-1, tadA) (ompH, oma87, ompA, plpB) (toxA) (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur); Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro;
 (ompH, oma87, ompA, plpB) (toxA) (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur); Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro;")
60
Fatores de virulência Cápsula e LPS Fímbrias e adesinas
Porção lipídica glicosídica (adaptado de FÁTIMA et al., 2005) Fatores de virulência Glicolipídeo Glicoproteína Porção lipídica Cápsula e LPS Fímbrias e adesinas Proteínas externas de membrana Toxina dermonecrótica (ptfA, pfhA, hsf-1, tadA) (ompH, oma87, ompA, plpB) (toxA) Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro; (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur);
 Fatores de virulência. Glicolipídeo. Glicoproteína. Porção. lipídica. Cápsula e LPS. Fímbrias e adesinas. Proteínas externas de membrana. Toxina dermonecrótica. (ptfA, pfhA, hsf-1, tadA) (ompH, oma87, ompA, plpB) (toxA) Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro; (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur);")
61
Fatores de virulência Cápsula e LPS Fímbrias e adesinas
Cu Zn Ligação dissulfeto Fatores de virulência Cápsula e LPS Fímbrias e adesinas Proteínas externas de membrana Toxina dermonecrótica (ptfA, pfhA, hsf-1, tadA) (ompH, oma87, ompA, plpB) (toxA) Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro; (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur);
 (ompH, oma87, ompA, plpB) (toxA) Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro; (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur);")
62
Complexo energético TonB
Transferrina Sideróforos Hemoglobina (adaptado de KREWULAK; VOGEL, 2008) Periplasma Fatores de virulência Citoplasma Cápsula e LPS Fímbrias e adesinas Proteínas externas de membrana Toxina dermonecrótica (ptfA, pfhA, hsf-1, tadA) (ompH, oma87, ompA, plpB) (toxA) Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro; Complexo energético TonB (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur);
 Periplasma. Fatores de virulência. Citoplasma. Cápsula e LPS. Fímbrias e adesinas. Proteínas externas de membrana. Toxina dermonecrótica. (ptfA, pfhA, hsf-1, tadA) (ompH, oma87, ompA, plpB) (toxA) Sialidases; Superóxido dismutase; Relacionados ao metabolismo do ferro; Complexo energético TonB. (nanH, nanB) (sodA, sodC) (hgbA, hgbB, exBD-tonB, fur);")
65
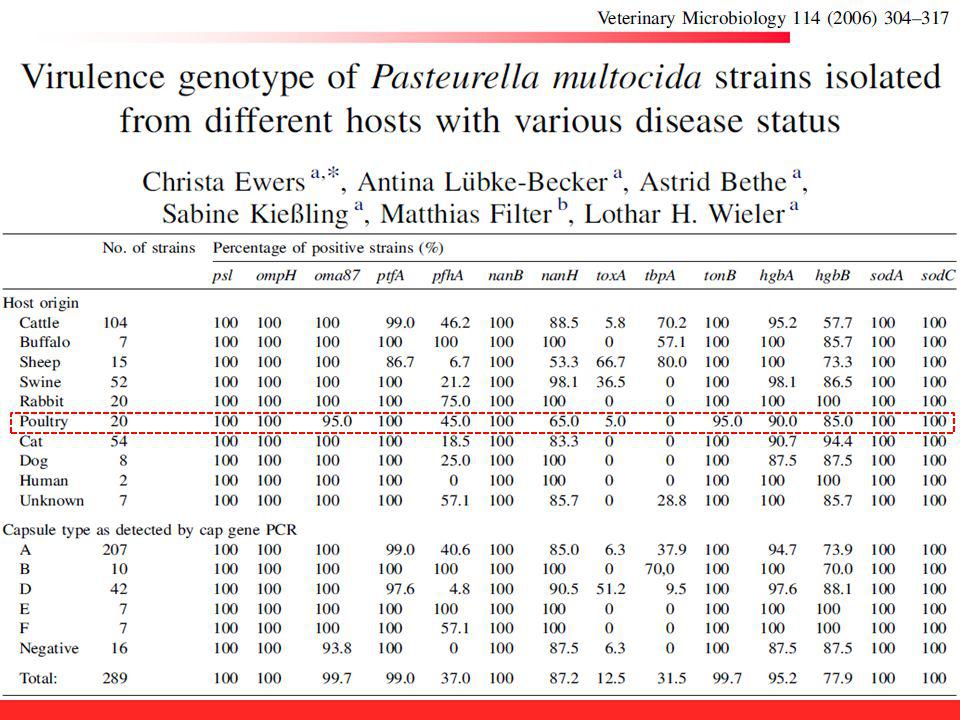
Tabela 5. Distribuição de 22 genes associados à virulência em Pasteurella multocida detectados por m-PCR conforme o hospedeiro de origem Processo ou enzima Gene Frequência absoluta e relativa (%) – cepas aviárias Frequência absoluta e relativa (%) – cepas suínas Proteínas de membrana externa ompH 96% 98% oma87 100% ompA 61% 95% plpB psl Metabolismo do ferro exbD-tonB fur hgbA 92% hgbB 93% 58% Sialidases nanH 85% nanB Superóxido dismutases sodA sodC Hyaluronic synthetase pmHAS 88% Toxina dermonecrótica toxA 0% 3% Adesinas ptfA pfhA 63% 53% tadD 38% hsf-1 50% 20% Submetido a Brazilian Journal of Microbiology (out.2014)
 – cepas aviárias. Frequência absoluta e relativa (%) – cepas suínas. Proteínas de membrana externa. ompH. 96% 98% oma % ompA. 61% 95% plpB. psl. Metabolismo do ferro. exbD-tonB. fur. hgbA. 92% hgbB. 93% 58% Sialidases. nanH. 85% nanB. Superóxido dismutases. sodA. sodC. Hyaluronic synthetase. pmHAS. 88% Toxina dermonecrótica. toxA. 0% 3% Adesinas. ptfA. pfhA. 63% 53% tadD. 38% hsf-1. 50% 20% Submetido a Brazilian Journal of Microbiology (out.2014)")
66
A maioria dos genes de virulência apresenta uma distribuição regular;
Alguns genes estão associados com a presença de pasteurelose (EWERS et al., 2006; BETHE et al., 2009; TANG et al., 2009) toxA+ tbpA+, pfhA+, hgbB+ Em aves não existem relatos de acordo com o status sanitário dos animais Alguns genes estão associados a determinados sorogrupos: Sorogrupo D toxA+, pmHAS-, hsf-1- pfhA+, pmHAS+, hsf-1- Sorogrupo A
 toxA+ tbpA+, pfhA+, hgbB+ Em aves não existem relatos de acordo com o status sanitário dos animais. Alguns genes estão associados a determinados sorogrupos: Sorogrupo D. toxA+, pmHAS-, hsf-1- pfhA+, pmHAS+, hsf-1- Sorogrupo A.")
67
Tabela 6. Frequência absoluta e relativa (%) dos genes associados à virulência detectados por PCR conforme os sorogrupos A e D de Pasteurella multocida Gene Sorogrupo A (gene hyaD-hyaC) (n=88) Sorogrupo D (gene dcbF) (n=6) ompH 85 (97%) 6 (100%) ompA 65 (74%) plpB 86 (98%) psl 5 (83%) exbD-tonB fur hgbA 84 (95%) hgbB 67 (76%) nanH 79 (90%) nanB 87 (99%) sodA sodC pmHAS 86 (98%)b 0 (0%)b toxA 0 (0%) 1 (17%)a ptfA pfhA 56 (64%)a 0 (0%)a tadD 53 (60%) hsf-1 28 (32%)b 6 (100%)a aem negrito: associação significativa (p<0.05) bem negrito: associação significativa (p<0.001) Submetido a Brazilian Journal of Microbiology (out.2014)
 (n=88) Sorogrupo D. (gene dcbF) (n=6) ompH. 85 (97%) 6 (100%) ompA. 65 (74%) plpB. 86 (98%) psl. 5 (83%) exbD-tonB. fur. hgbA. 84 (95%) hgbB. 67 (76%) nanH. 79 (90%) nanB. 87 (99%) sodA. sodC. pmHAS. 86 (98%)b. 0 (0%)b. toxA. 0 (0%) 1 (17%)a. ptfA. pfhA. 56 (64%)a. 0 (0%)a. tadD. 53 (60%) hsf (32%)b. 6 (100%)a. aem negrito: associação significativa (p<0.05) bem negrito: associação significativa (p<0.001) Submetido a Brazilian Journal of Microbiology (out.2014)")
68
Tabela 7. Percentual de genes associados aos pares entre 96 cepas de Pasteurella multocida analisadas ompH toxA ptfA nanH exbD-tonB sodA pfhA hgbA sodC nanB hgbB oma87 hsf-1 fur pmHAS psI ompA plpB tadA 100 1 97 93* 100 90 100* 98 99 58 57 60 59 96 99* 78 80 77 82* 36* 38 33* 36 37 35* 42 98* 92* 89 87 78* psl 77* 76 76* 44 75 63* 74 68* 39** 73 100** 94 tadD 57* 56 48* 55 45** 28* 59* 62* 74* *em negrito: indica associação significativa (p<0.05) **em negrito: indica associação significatava (p<0.001) Submetido a Brazilian Journal of Microbiology (out.2014)
 **em negrito: indica associação significatava (p<0.001) Submetido a Brazilian Journal of Microbiology (out.2014)")
69
Tabela 7. Percentual de genes associados aos pares entre 96 cepas de Pasteurella multocida analisadas ompH toxA ptfA nanH exbD-tonB sodA pfhA hgbA sodC nanB hgbB oma87 hsf-1 fur pmHAS psI ompA plpB tadA 100 1 97 93* 100 90 100* 98 99 58 57 60 59 96 99* 78 80 77 82* 36* 38 33* 36 37 35* 42 98* 92* 89 87 78* psl 77* 76 76* 44 75 63* 74 68* 39** 73 100** 94 tadD 57* 56 48* 55 45** 28* 59* 62* 74* *em negrito: indica associação significativa (p<0.05) **em negrito: indica associação significatava (p<0.001) Submetido a Brazilian Journal of Microbiology (out.2014) Houve associação significativa na combinação da presença dos genes de virulência em 45 situações.
 **em negrito: indica associação significatava (p<0.001) Submetido a Brazilian Journal of Microbiology (out.2014) Houve associação significativa na combinação da presença dos genes de virulência em 45 situações.")
70
Habitante normal das vias aéreas
(MUHAIRWA et al., 2000; SAMUEL et al., 2007; WILKIE et al., 2012) ? ? ? cólera aviária saúde animal ?
 cólera aviária. saúde animal.")
71
Habitante normal das vias aéreas
(MUHAIRWA et al., 2000; SAMUEL et al., 2007; WILKIE et al., 2012) ? ? ? cólera aviária saúde animal ? A bactéria é patogênica?
 cólera aviária. saúde animal. A bactéria é patogênica")
72
? multocida septica A1:3 colônia iridescente hgbA+ Β-hemólise
BIOVAR 3 BIOVAR 13 multocida septica gallicida BIOVAR 2 A:1 A1:3 sorogrupo D cápsula tipo F colônia iridescente pfhA+ hgbA+ toxA+ hgbB+ Β-hemólise fur nanH

73
Genoma Pm70 Dendrograma com a relação de cepas de P. multocida conforme o perfil de virulência Alinhamento e comparação da sequência de aminoácidos da proteína OmpH da cepa P52 com diferentes sorotipos de P. multocida raros variáveis quanto ao padrão de avaliação Utilização da inoculação de animais para determinar a patogenicidade dos isolados Lei (08/10/08) Procedimentos para o uso científico de animais (Ibrahim et al., 1998; Shivachandra et al., 2006; Mohamed et al.,2012)
 Procedimentos para o uso científico de animais. (Ibrahim et al., 1998; Shivachandra et al., 2006; Mohamed et al.,2012)")
74
Estabelecimento de um índice de patogenicidade em pintos de corte de um dia de idade para cepas de Pasteurella multocida isoladas de de aves e de suínos. (Pilatti, 2014) estabelecer um modelo de classificação da patogenicidade de P. multocida a partir da inoculação de pintos de corte. Parâmetros avaliados tempo de morte (dias): observação das aves por 07 dias 2. presença de lesões macroscópicas A B C D E F A) aerossaculite B) Lesão local aplic. C) onfalite D) pericardite E) perihepatite F) peritonite
 estabelecer um modelo de classificação da patogenicidade de P. multocida a partir da inoculação de pintos de corte. Parâmetros avaliados. tempo de morte (dias): observação das aves por 07 dias. 2. presença de lesões macroscópicas. A. B. C. D. E. F. A) aerossaculite. B) Lesão local aplic. C) onfalite. D) pericardite. E) perihepatite. F) peritonite.")
75
Somatório do Escore de Lesões (EL)
Cálculo do Índice de Patogenicidade Individual (IPI) para cada animal somatório escore de lesões *IPI = TM x 5 + (PC + PH + PT + ASS + CL + ONF) *TM: Tempo de Morte; PC: Pericardite; PH: Perihepatite; PT: Peritonite; ASS: Aerossaculite; LA: local de aplicação; ONF: Onfalite; Fator de bonificação de sobrevivência: 0,1428 (Souza, 2010) Tabela 8. Relação entre o tempo de morte ou o número de lesões e o respectivo valor atribuído Dia de Morte TM TM x 5 Somatório do Escore de Lesões (EL) N° de Lesões Valor 1 5 6 5,000 2 0,8572 4,286 4,165 3 0,7144 3,571 4 3,333 0,571 2,857 2,500 0,4284 2,143 1,667 0,2857 1,428 0,833 7 0,1428 0,714 0,000 7* *** Cálculo do Índice de Patogenicidade Individual (IP) para a cepa inoculada IP = ∑(IPI) N 10 aves p/ cada cepa (Pilatti, 2014)
 para cada animal. somatório escore de lesões. *IPI = TM x 5 + (PC + PH + PT + ASS + CL + ONF) *TM: Tempo de Morte; PC: Pericardite; PH: Perihepatite; PT: Peritonite; ASS: Aerossaculite; LA: local de aplicação; ONF: Onfalite; Fator de bonificação de sobrevivência: 0,1428. (Souza, 2010) Tabela 8. Relação entre o tempo de morte ou o número de lesões e o respectivo valor atribuído. Dia de Morte. TM. TM x 5. Somatório do Escore de Lesões (EL) N° de Lesões. Valor , , ,286. 4, , , ,333. 0,571. 2,857. 2,500. 0, ,143. 1,667. 0, ,428. 0, , ,714. 0,000. 7* *** Cálculo do Índice de Patogenicidade Individual (IP) para a cepa inoculada. IP = ∑(IPI) N. 10 aves p/ cada cepa. (Pilatti, 2014)")
76
Classificação de Patogenicidade
Tabela 9. Valores dos Índices de Patogenicidade (IP) das 96 cepas de Pasteurella multocida inoculadas em frangos de corte com 01 dia idade Cepa IP 1 10 17 33 7,80 49 6,11 65 4,44 81 2,25 2 18 9,18 34 7,75 50 5,81 66 4,43 82 2,17 3 19 9,13 35 7,69 51 5,63 67 4,39 83 2,1 4 20 8,93 36 52 5,46 68 3,95 84 5 21 8,87 37 7,5 53 5,44 69 3,89 85 1,75 6 22 8,69 38 7,57 54 5,26 70 3,79 86 1,67 7 23 8,6 39 7,44 55 5,24 71 3,60 87 1,58 8 24 8,51 40 7,25 56 5,17 72 3,5 88 1,42 9 25 8,44 41 7,20 57 5,07 73 3,31 89 1,33 26 8,27 42 6,93 58 4,93 74 3,27 90 11 27 8,26 43 6,77 59 4,89 75 3,12 91 12 28 8,13 44 6,65 60 4,88 76 2,99 92 13 29 8,06 45 6,56 61 4,80 77 2,6 93 1,25 14 30 7,93 46 6,43 62 4,73 78 2,5 94 1,17 15 31 7,90 47 6,42 63 4,69 79 2,42 95 1,08 16 32 7,82 48 6,31 64 4,56 80 2,29 96 0,17 Tabela 10. Mediana dos valores dos Índices de Patogenicidade (IP) de acordo com a distribuição das cepas de Pasteurella multocida nos grupos de patogenicidade Classificação de Patogenicidade N° de Amostras Mediana ± DP Alta (8-10) 37 9,13 ± 0,953 ᵃ Intermediária (4-7) 35 5,24 ± 1,159 ᵇ Baixa (0-3) 24 1,75 ± 0,837 ᶜ (Pilatti, 2014) Letras diferentes na mesma coluna representam diferença estatística significativa (p<0,01) Kruskall-Wallis test
 das 96 cepas de Pasteurella multocida inoculadas em frangos de corte com 01 dia idade. Cepa. IP , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , ,17. Tabela 10. Mediana dos valores dos Índices de Patogenicidade (IP) de acordo com a distribuição das cepas de Pasteurella multocida nos grupos de patogenicidade. Classificação de Patogenicidade. N° de Amostras. Mediana ± DP. Alta (8-10) 37. 9,13 ± 0,953 ᵃ. Intermediária (4-7) 35. 5,24 ± 1,159 ᵇ. Baixa (0-3) 24. 1,75 ± 0,837 ᶜ. (Pilatti, 2014) Letras diferentes na mesma coluna representam diferença estatística significativa (p<0,01) Kruskall-Wallis test.")
77
Redes Neurais Artificiais
Explicar, simular e predizer os resultados zootécnicos da cadeia avícola; Gerenciamento de reprodutoras pesadas Gerenciamento de frangos de corte Gerenciamento de matadouros-frigoríficos de aves Guahyba, 2001 Realli, 2004 Pinto, 2006 Spohr, 2010 Gerenciamento da cadeia avícola Explicar, simular e predizer os resultados em temas sanitários da cadeia avícola; Moraes, 2008 Carvalho, 2013 Rocha, 2006; Tejkowski, 2013 Salle, 2009; Da Rocha, 2012 Avaliação da depleção linfocitária da Bursa de Fabricius Avaliação da depleção linfocitária do timo Classificação da patogenicidade de Escherichia coli Classificação da resistência antimicrobiana de Escherichia coli

78
Redes Neurais Artificiais
Simula em computadores o funcionamento do cérebro humano implementando modelos matemáticos que se assemelhem às estruturas neurais biológicas; (FERNEDA, 2006) RNAs calculam funções matemáticas normalmente não lineares e estão dispostas em uma ou mais camadas interligadas por um grande número de conexões associados a determinados pesos conforme as informações de entrada recebidas. (BRAGA et al., 2000; LUDWIG & COSTA, 2007) dendritos sinapses
 RNAs calculam funções matemáticas normalmente não lineares e estão dispostas em uma ou mais camadas interligadas por um grande número de conexões associados a determinados pesos conforme as informações de entrada recebidas. (BRAGA et al., 2000; LUDWIG & COSTA, 2007) dendritos. sinapses.")
79
Redes Neurais Artificiais
Camada de entrada Camada de saída Presença de 22 genes de virulência Classificação do índice de patogenicidade das cepas Alta Intermediária Baixa

80
Programa Neuroshell Classifier 2.1

81
Alternativa para determinação da patogenicidade sem a utilização de modelos animais
Programa Neuroshell Classifier 2.1

82
Relação epidemiológica de um grupo de genes
Amostras isoladas de animais sadios e doentes Amostras de diferentes origens CA na forma aguda CA na forma crônica Relação epidemiológica de um grupo de genes Os genes toxA, tbpA e pfhA: marcadores do potencial patogênico em outras espécies (DAVIES et al., 2004; BETHE et al., 2009) Obtenção de amostras isoladas em outros hospedeiros (ruminantes, suínos, pets); Possibilidade de transmissão horizontal de genes de virulência (DAVIES et al., 2004; BETHE et al., 2009) Comparação perfis genéticos e tipificação molecular (ZAGLIC et al., 2005; SUBAAHARAN et al., 2010)
 Obtenção de amostras isoladas em outros hospedeiros (ruminantes, suínos, pets); Possibilidade de transmissão horizontal de genes de virulência. (DAVIES et al., 2004; BETHE et al., 2009) Comparação perfis genéticos e tipificação molecular. (ZAGLIC et al., 2005; SUBAAHARAN et al., 2010)")
83
Obrigado pela atenção.

Apresentações semelhantes






. Nenhuns direitos reservados, excepto para fins comerciais. Por favor, não coloque.>")














