Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Design, síntese total e superexpressão funcional do gene lip1 de Candida rugosa codificando para uma grande lipase industrial S. BROCCA, C. SCHMIDT-DANNERT, M. LOTTI, L. ALBERGHINA and R. D. SCHMID, 1998 Amanda F. Sarmento Nathani C. B. Denardi Rafael L. Farinha Disciplina: Engenharia de proteínas Prof. Dr. Jesus A. Ferro Jaboticabal Setembro de 2013

2
Design, síntese total e superexpressão funcional do gene lip1 de Candida rugosa codificando para uma grande lipase industrial S. BROCCA, C. SCHMIDT-DANNERT, M. LOTTI, L. ALBERGHINA and R. D. SCHMID, 1998 Amanda F. Sarmento Nathani C. B. Denardi Rafael L. Farinha Disciplina: Engenharia de proteínas Prof. Dr. Jesus A. Ferro Jaboticabal Setembro de 2013

3
Introdução Lipases As lipases (Triacilglicerol acil-hidrolases EC ) são enzimas que catalisam a hidrólise de triacilglicerídeos na interface água/meio orgânico. In vitro são enzimas versáteis devido a sua capacidade de catalisar a hidrólise e a síntese de uma grande variedade de ésteres, ou a resolução de misturas racémicas De maneira geral, as lipases não requerem cofatores, atuam em ampla faixa de pH, são estáveis à altas temperaturas, que fazem com que sejam altamente aplicáveis em processos industriais (VILLENEUVE et al., 2000; HASAN, SHAH, HAMEED, 2006).
 são enzimas que catalisam a hidrólise de triacilglicerídeos na interface água/meio orgânico. In vitro são enzimas versáteis devido a sua capacidade de catalisar a hidrólise e a síntese de uma grande variedade de ésteres, ou a resolução de misturas racémicas. De maneira geral, as lipases não requerem cofatores, atuam em ampla faixa de pH, são estáveis à altas temperaturas, que fazem com que sejam altamente aplicáveis em processos industriais (VILLENEUVE et al., 2000; HASAN, SHAH, HAMEED, 2006).")
4
Introdução Muitos processos tecnológicos utilizam catalisadores biológicos nas sequências de conversão química. As principais fontes de obtenção de lipases para aplicação industrial têm sido os microorganismos. Tanto micro-organismos eucariotos (leveduras) como procariotos (bactérias), são produtores de lipases e suas propriedades variam de acordo com a procedência. O uso de enzimas nas indústrias possibilita o desenvolvimento de processos tecnológicos tão eficientes quanto aos realizados pela natureza (HASAN; SHAH; HAMEED, 2006) e sem causar riscos ambientais.
 como procariotos (bactérias), são produtores de lipases e suas propriedades variam de acordo com a procedência. O uso de enzimas nas indústrias possibilita o desenvolvimento de processos tecnológicos tão eficientes quanto aos realizados pela natureza (HASAN; SHAH; HAMEED, 2006) e sem causar riscos ambientais.")
5
Introdução Algumas aplicações da lipase: Indústrias de detergentes;
Medicamentos; Alimentos (panificação, queijos e chás); Têxteis; Cosméticos; Biodiesel; - Tratamento de efluentes;
; Têxteis; Cosméticos; Biodiesel; - Tratamento de efluentes;")
6
Introdução Candida rugosa
- Trata-se de uma levedura do gênero Candida sp. - As lipases de Candida rugosa estão entre as lipases comercias mais frequentemente utilizadas na hidrólise e a síntese de uma gama de ésteres de interesse comercial (Kotting & Eibl, 1994; Vulfson, 1994). - Na maioria das aplicações biocatalíticas preparações de enzima em bruto, obtidas com TCA (Ácido tricloroacético) do sobrenadante da cultura, são aplicadas.
. - Na maioria das aplicações biocatalíticas preparações de enzima em bruto, obtidas com TCA (Ácido tricloroacético) do sobrenadante da cultura, são aplicadas.")
7
Introdução - Uma alta homologia de sequência (entre 60-70%) entre vários genes de lipase que foram clonados a partir de C. rugosa. - Diferenças na glicolisação destas lipases podem contribuir para esta heterogeneidade. - Várias isoformas de lipase foram isoladas a partir de preparação de enzimas comerciais , sendo mostrado que o produto do gene lip1 é o constituinte principal. - Pelo fato das isoformas poderem diferir nos seus desempenhos catalíticos e suas propriedades poderem ainda ser afetadas pelos processos de purificação, pensou-se que a clonagem e expressão do gene lip1 seria a abordagem mais adequada para produção e caracterização de lipase pura de C. rugosa.

8
Introdução - C. rugosa dimórfica obedece a uma utilização de códons em que o não-canônico tripleto CUG, um códon universal para leucina, é lido com serina. - Na maioria das espécies de Candida sp., filogeneticamente relacionados , CUG é extremamente raro, com notável exceção de C. rugosa , onde responde por cerca de 40% do total de códons de serina. - No gene Lip1, 20 dos seus 47 resíduos de serina, incluindo a Ser209 catalítica, são codificados por tripletos CUG. Como consequência, a expressão heteróloga de Lip1 em Saccharomyces cerevisiae resulta em uma lipase inativa. A troca de vários , ou mesmo a maioria, códons CUG por tripletos universais de serina (UCN, AGY) é necessária para expressão em hospedeiro heterólogo.
 é necessária para expressão em hospedeiro heterólogo.")
9
Introdução Neste trabalho foram investigadas duas abordagens para a expressão da proteína funcional de Lip1 : - Mutagênese de um gene Lip1 natural e a sua expressão em S. cerevisiae; - Síntese química enzimática do mesmo gene. Três genes sintéticos recombinantes (slip1) foram construídos diferindo na sequência líder . Os genes recombinantes foram expressos em S. cerevisiae e Picha pastoris.
 foram construídos diferindo na sequência líder . Os genes recombinantes foram expressos em S. cerevisiae e Picha pastoris.")
10
Introdução Picha pastoris - É uma espécie de levedura metilotrófica.
Como em S. cerevisiae , a secreção pelas células requer a presença de uma sequência de sinal fundido com a proteína expressa. Propriedades físico-químicas e catalíticas da lipase recombinante produzidas por P. pastoris, foram comparadas com as CRLs comercias, revelando que são idênticas.

11
Mutagênese sítio-dirigida do gene da lipase de C. rugosa
Cinco membros, pertencentes à família de multigenes de codificação para as lipases de C. rugosa. Utilização da sequência Lip1 Melhor caracterizado; Estrutura cristalográfica resolvida; Codifica a principal isoforma da lipase; Lip1 contém sequência genômica com uma única ORF de 1647 pb, o que corresponde a uma proteína de 549 resíduos com uma extensão N-terminal de 15 resíduos hidrofóbicos, que codificam um peptídeo sinal .

12
Mutagênese sítio-dirigida do gene da lipase de C. rugosa
A proteína madura Lip1 contém 46 resíduos de Serina, 19 delas codificadas por tripletos CTG. A primeira tentativa de superar os problemas que ocorrem na expressão heteróloga foi a substituição, por mutagênese sítio-dirigida dos códons CTG por outros códons universais para Serina. Hierarquia dos resíduos de Ser, funcionalmente e estruturalmente mais importante a ser mutado.

13
Mutantes de Serina Com base no alinhamento com outras Serinas-hidrolases e sobre a estrutura 3D da enzima são selecionadas oito Ser como alvo para mutagênese. - Quatro genes mutantes, contendo um Nº crescente de resíduos de Ser restaurados.

14
Mutagênese sítio-dirigida do gene da lipase de C. rugosa
Genes mutantes foram clonados no vetor pEMBLyex4, contendo a sequência de indução. Células de leveduras recombinantes, cultivas sob condições de indução, acumulam intracelularmente produto inativo do gene Lip1 a um nível de 10-20mg/L da cultura. Embora a análise da glicolisação de proteínas recombinantes tenha fornecido evidências de N-glicolisação, como no caso de Lip1 do tipo selvagem, sua secreção falhou sugerindo dificuldades no nível pós-transcrional.

15
Construção de um material sintético, com códons optimizados da lipase da C. rugosa

16
Expressão da lipase recombinante em P. pastoris e S. cerevisiae
Gene sintético nl-slip 1 PCR PCR pp-slip 1 p-slip 1 Vetores pPIC-nl-slip1 pPIC-pp-slip1 pPIC-p-slip1 pY-nl-slip1 pY-pp-slip1 pY-p-slip1 pPICZαB pYES2 P. pastoris S. cerevisiae

17
Expressão dos genes sintéticos da Lip1 em P.pastoris
Pichia Pastoris e Metanol -> Fonte de Carbono AOX regula Metanol Secreção evidenciada por “anéis” nas placas de tributirina-metanol. pPIC-pp-slip1 e pPIC-p-slip1 -> Atividade Overnight pPIC-NL-slip1 ->Atividade (48h-72h) pPICZ-alfa-B -> Sem atividade
 pPICZ-alfa-B -> Sem atividade.")
18
Expressão dos genes sintéticos da Lip1 em P.pastoris
Seleção dos melhores clones através da formação de anéis. Cultivo Metanol 20mL (Fonte de Carbono) Indução: 48h Palmitato de p-nitrofenil -> Sobrenadante (Multiplas Inserções Genômicas) BMMY 200 ml (Meio Rico Padrão) Indução: 5dias
 Indução: 48h. Palmitato de p-nitrofenil -> Sobrenadante. (Multiplas Inserções Genômicas) BMMY 200 ml (Meio Rico Padrão) Indução: 5dias.")
19
Expressão dos genes sintéticos da Lip1 em P.pastoris
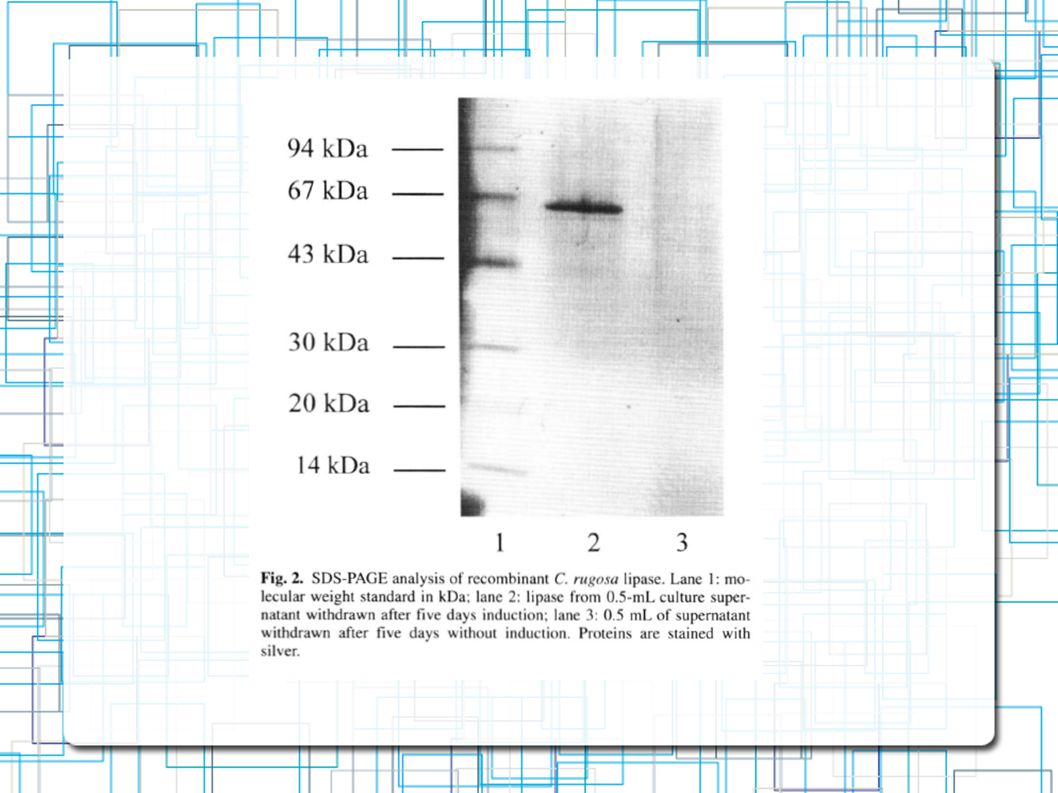
Atividade da Lipase: pPIC-p-slip1 pPIC-pp-slip1 SDS-PAGE Única Banda de 60 Kda Peso molecular já era esperado para a Lip1 Nenhuma outra proteína foi secretada pela P.Pastoris 85 U/ml

21
Expressão dos genes sintéticos da Lip1 em P.pastoris
Outros testes executados Western-Blott Sequênciamento Amino-Terminal Acúmulo de Lipase recombinante no meio Intracelular -> Seq. Pré-Pro Sinalizadora do Fator Alfa (pPIC-pp-slip1) -> Apenas Expressão Não houve acúmulo -> Seq. Sinalizadora Natural -> Apenas Secreção Seq. Líder sinaliza a secreção na P.pastoris, evitando o acúmulo. Seq. Pré-Sinalizadora do Fator Alfa (pPIC-p-slip1) -> Secreção E expressão
 -> Apenas Expressão. Não houve acúmulo -> Seq. Sinalizadora Natural -> Apenas Secreção. Seq. Líder sinaliza a secreção na P.pastoris, evitando o acúmulo. Seq. Pré-Sinalizadora do Fator Alfa (pPIC-p-slip1) -> Secreção E expressão.")
22
Expressão dos genes sintéticos da Lip1 em P.pastoris
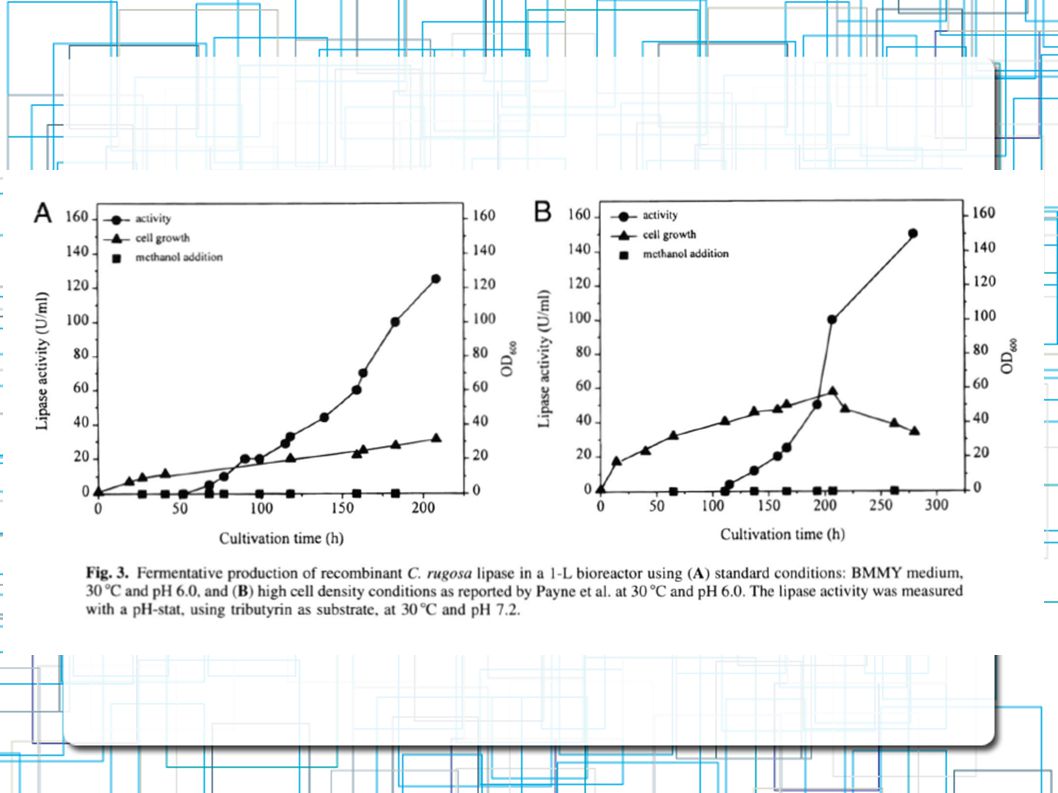
Clone pPIC-pp-slip1 com o maior nível de secreção de lipase Fermentação com Biorreator BMMY pH 6 208h Para melhorar ainda... Fermentação Celular de Alta Densidade 280h 125 U/ml 150 U/ml

24
Expressão dos genes sintéticos da Lip1 em P.pastoris
Produtividade de U/L por hora. Lipase Nativa na C.rugosa Ácido Oleico X Metanol (Fonte de Carbono_ Produção de 250 U/L por hora. P.pastoris com lip1 tem o dobro da produtividade

25
Expressão dos genes sintéticos da Lip1 em S.cerevisiae
Transformantes obtidos por vetores: pY-nl-slip1 pY-pp-lip1 pY-p-slip1 Atividade lipásica Placas de Tributirina-Galactose Indução: 24h 30ºC Transformante obtido por vetor: Pyes2 -> Não teve atividade

26
Expressão dos genes sintéticos da Lip1 em S.cerevisiae
Os melhores clones foram cultivados: Erlenmeyes 2-L 200mL meio de Glicose Mínimo Até atingir Densidade Óptica 600 5 dias de indução -> Meio mínimo Galactose Atividade dos sobrenadantes: 5-7U/mL (vetores pY-pp-slip1, pY-p-slip1, pY-nl-slip1) 12-17x menos efetivo que na P.pastoris em cultivo de pequena escala
 12-17x menos efetivo que na P.pastoris em cultivo de pequena escala.")
27
Expressão dos genes sintéticos da Lip1 em S.cerevisiae
Para o gene natural e modificado -> Completa falha em relação a lipase Sem Acúmulo Sem Secreção

28
Caracterização da Lipase Recombinante
Elétroforese de poliacrilamida do plasmídio pPIC-p-slip1 da P.pastoris -> Única banda de 60 kDA. Lip1 Nativa e suas Isoformas A sequencia de aminoacidos da Lip1 contem três sítios potenciais de N-glicosilação nas posições 291, 314 e 351 Após a Deglicosilação: Perda de 3kDa -> 5% do peso eram carboídratos Nas Lipases não recombinantes, o peso é de cerca de 3,6-8% de carboídratos Realizada antes e depois da Desnaturação Peso molecular igual

29
Caracterização da Lipase Recombinante
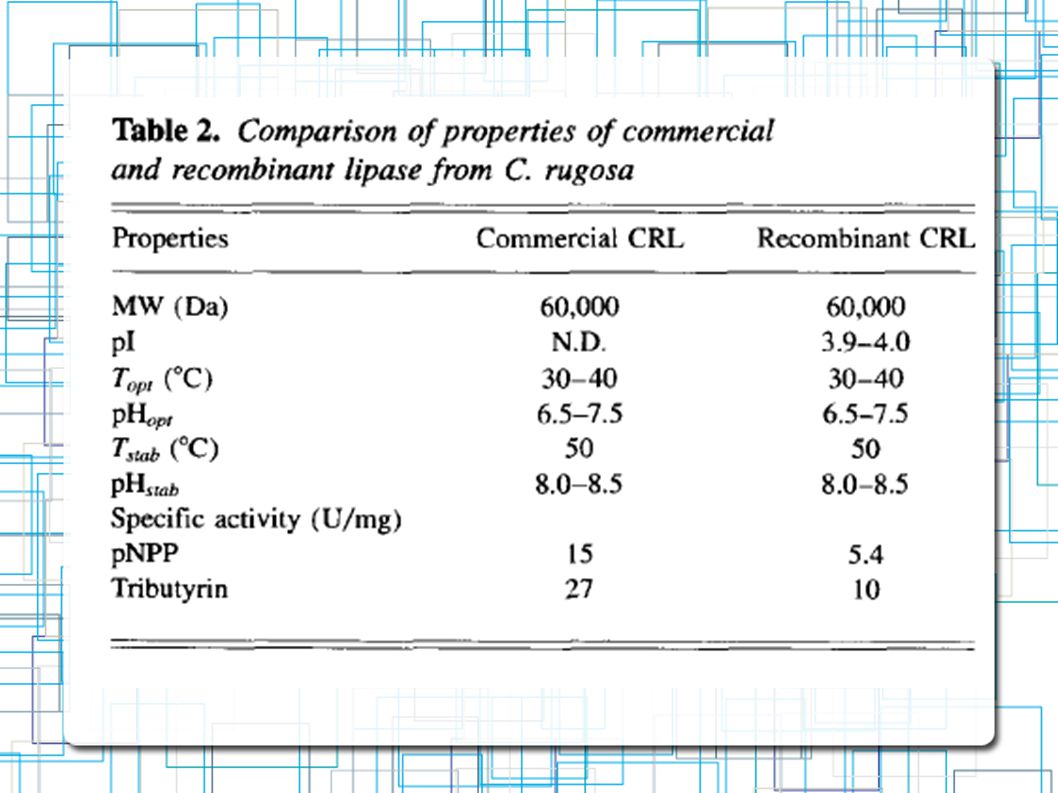
C.rugosa comercial Lip1 ativa em 30-40ºC pH 7 (Efetividade reduzida em 60% se elevado a pH 8) Dois pI pouco diferentes (3,9 e 4)
 Dois pI pouco diferentes (3,9 e 4)")
31
Caracterização da Lipase Recombinante
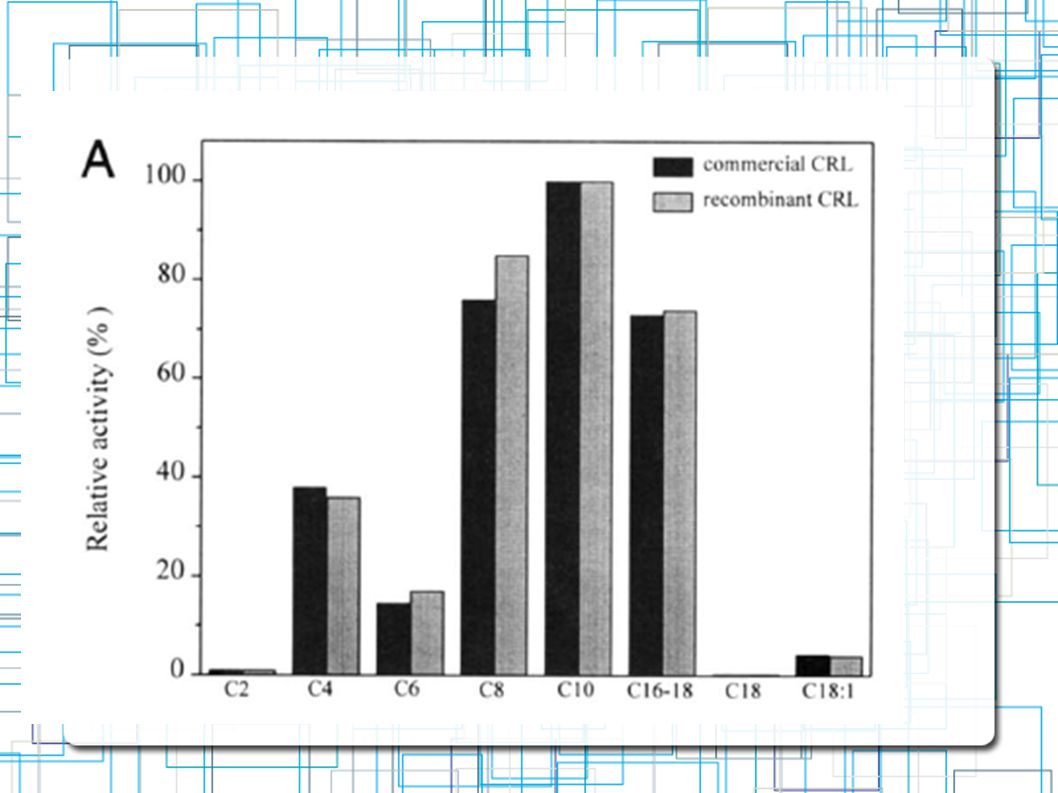
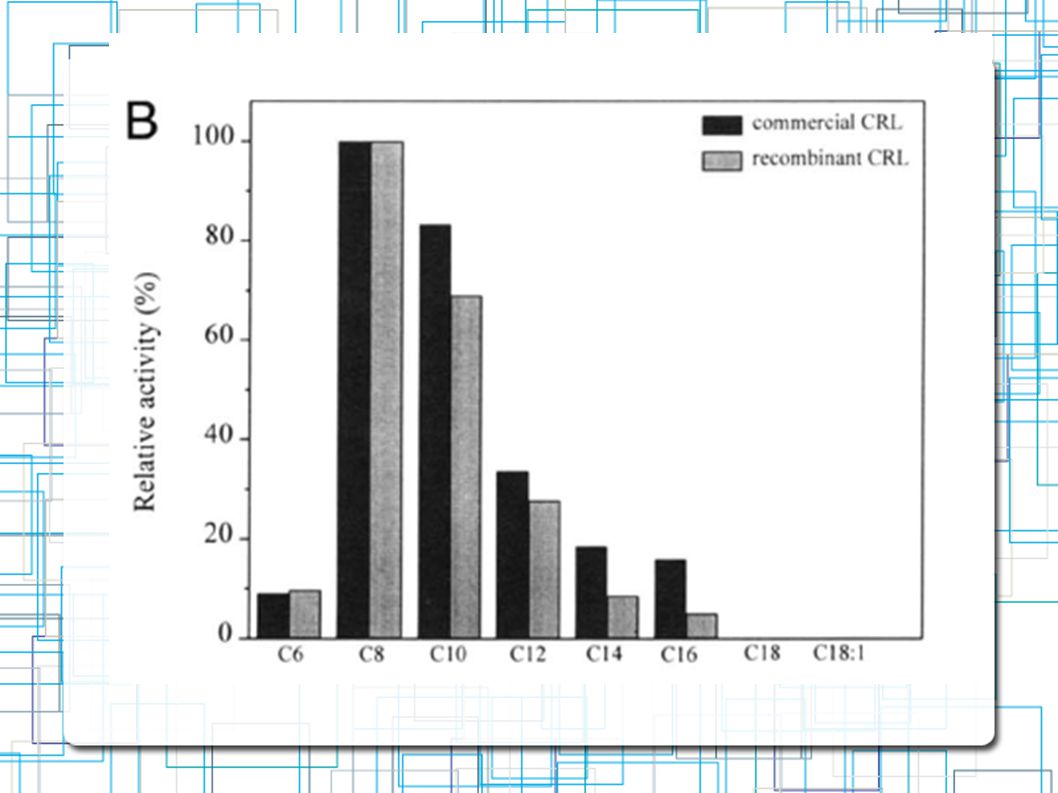
Comparação da Especifidade de substratos entre lip1 e lipase da C.rugosa Triglicerideos com variação em tamanho de cadeias de grupos acyl Atividade Lipolítica Alta Tricaproina (C8) Tricaprina (C10) Atividade Lipolítica Baixa Trioleina (C18) Atividade HIDROlítica Alta Mantega de Cacau(C16-18)
 Tricaprina (C10) Atividade Lipolítica Baixa. Trioleina (C18) Atividade HIDROlítica Alta. Mantega de Cacau(C16-18)")
33
Caracterização da Lipase Recombinante
Comparação da Especifidade de substratos entre lip1 e lipase da C.rugosa Ésteres Metílicos com variação em tamanho de cadeias de grupos acyl (C6-C22) Após a atividade da lipase, os acidos graxos foram quantificados por Cromatografia em Gás Atividade Lipolítica Alta C8 - C10 Atividade Lipolítica NULA C18 – C22
 Após a atividade da lipase, os acidos graxos foram quantificados por Cromatografia em Gás. Atividade Lipolítica Alta. C8 - C10. Atividade Lipolítica NULA. C18 – C22.")
35
Caracterização da Lipase Recombinante
Comparação da Especifidade de substratos entre lip1 e lipase da C.rugosa Ésteres Metílicos com variação em tamanho de cadeias de grupos acyl(C6-C22) Após a atividade da lipase, os acidos graxos foram quantificados por Cromatografia em Gás Atividade Lipolítica Alta C8 - C10 Atividade Lipolítica NULA C18 – C22
 Após a atividade da lipase, os acidos graxos foram quantificados por Cromatografia em Gás. Atividade Lipolítica Alta. C8 - C10. Atividade Lipolítica NULA. C18 – C22.")
36
Caracterização da Lipase Recombinante
Propriedades físíco-quimicas e catalíticas da Lip1 e CRL coincidem Lip1 em P.pastoris -> Mais funcionalmente Ativa

37
Protocolo Experimental

38
Cepas, plasmídeos e meios
hospedeiro para amplificação do plasmídeo E. coli: DH5a Plasmídeos: pUC19 e PCYTEXP1 PEMBLyex4 pYES (Invitrogen) pPICZαB (Invitrogen) P. pastoris GS115 (Invitrogen) S. cerevisiae INVSC2 (Invitrogen) S. cerevisae X4004.3A Expressão da lipase recombinante clonagem expressão
 pPICZαB (Invitrogen) P. pastoris GS115. (Invitrogen) S. cerevisiae INVSC2 (Invitrogen) S. cerevisae X4004.3A. Expressão da lipase recombinante. clonagem. expressão.")
39
Cepas, plasmídeos e meios
P. pastoris cultivada em frasco de agitação a 30ºC Meio rico padrão S. cerevisiae cultivada a 30ºC 0,67% (w/v) do meio mínimo: Base Nitrogenada de levedura(YNB) + aminoácido apropriado de 50mg/mL e 2% glicose ou 2% galactose como fonte de carbono PUC19 pCYTEXP1 E. coli cultivada a 37ºC Meio LB (Luria-Bertani) 100ug/mL de ampicilina ou 25 ug/mL de zeocina seleção dos clones transformados pPICZαB 1% de glicerol (BMGY) ou 0,5 de metanol (BMMY) indução
 do meio mínimo: Base Nitrogenada de levedura(YNB) + aminoácido apropriado de 50mg/mL e 2% glicose ou 2% galactose como fonte de carbono. PUC19. pCYTEXP1. E. coli. cultivada a 37ºC. Meio LB (Luria-Bertani) 100ug/mL de ampicilina. ou. 25 ug/mL de zeocina. seleção. dos clones. transformados. pPICZαB. 1% de glicerol (BMGY) ou. 0,5 de metanol (BMMY) indução.")
40
Cepas, plasmídeos e meios
Para manter as culturas de leveduras foi usado o meio YEPD (1% de extrato de levedura, 2% de peptona e 2% de dextrose) Para selecionar os transformantes de P. pastoris foram utilizadas placas contendo zeocina (100mg/mL) e meio YEPS (YEP + 2% de sorbitol)
 Para selecionar os transformantes de P. pastoris foram utilizadas placas contendo zeocina (100mg/mL) e meio YEPS (YEP + 2% de sorbitol)")
41
Cada cassete foi sintetizado em uma etapa
Design do gene, síntese do cassete e montagem do códon do gene otimizado Gene foi dividido em 4 fragmentos de ca. 400pb cada clonados separadamente 4 subcontruções resultantes: Após remoção de mutações indesejáveis Cassete I 4 oligonucleotídeos Cassete II Cassete III 6 oligonucleotídeos Cassete IV Cada cassete foi sintetizado em uma etapa corte abrupto Vetor de clonagem pUC19 digerido com Smal pUC-I pUC-II clonados em pUC19 pUC-III pUC-IV

42
Design do gene sintético
Cassete I Cassetes II a IV 4 oligonucleotídeos 6 oligonucleotídeo gene Lip1

43
Design do gene sintético
O gene completo da lipase sintética foi montade em pUC19 Posteriormente, o fragmento XmaI-EcoRI de pUC-(II-III) foi inserido no pUC-I resultando em pUC-(I-III) O fragmento de EcoRI-EcoRI da pUC-IV foi clonado em pUC-(I-III) Fragmento de XmaI/blunt-SalI da pUC-II estava ligado na pUC-III anteriormente linearizada com pUCII, pUC III PstI/Iblunt SalI XmaI e EcoRI cortado com EcoRI Vetor de clonagem corretamente montado pUC-(I-IV) Identificado e caracterizado pela análise de enzimas de restrição Sequência inteira foi reconfirmada pelo sequenciamento automatizado
 foi inserido no pUC-I resultando em pUC-(I-III) O fragmento de EcoRI-EcoRI da pUC-IV foi clonado em pUC-(I-III) Fragmento de XmaI/blunt-SalI. da pUC-II. estava ligado na pUC-III. anteriormente linearizada com. pUCII, pUC III. PstI/Iblunt. SalI. XmaI e EcoRI. cortado. com EcoRI. Vetor de clonagem. corretamente. montado pUC-(I-IV) Identificado e caracterizado pela análise de enzimas de restrição. Sequência inteira foi reconfirmada pelo sequenciamento automatizado.")
44
Construção dos vetores de expressão Lip1
Gene sintético Lip1 (slip) foi subclonado passo intermediário para a fusão: Fragmento BamHI de pUC(I-IV) estava ligado em um vetor linear de BamHI → pCYTEXPI, originando pCY-slip1 vetor pCYTEXP1 sequência líder do fatorα de S. cerevisiae gene vetor pCYTEXPI pCY-slip1 BamHI
 foi subclonado. passo intermediário para a fusão: Fragmento BamHI de pUC(I-IV) estava ligado em um vetor linear de BamHI → pCYTEXPI, originando pCY-slip1. vetor pCYTEXP1. sequência líder. do fatorα de. S. cerevisiae. gene. vetor. pCYTEXPI. pCY-slip1. BamHI.")
45
Construção de vetores de expressão de Lip1
PCR Sequência do fator alfa presente no vetor de expressão de P. pastoris (pPICZαB) Sequência que codifica a lipase Sequência que codifica a lipase Sequência que codifica a lipase primer complementar a sequência pPICZαB O produto obtido da PCR foi clonado em SphI/blunt linearizado com pCYTEXPI, resultando em pCY-prepo-fator-α O fragmento BglI de pCY-slip1, contendo toda a forma madura do gene sintético da lipase, foi inserido no plasmídeo digerido (pCyprepo-fator-α), restaurando assim o gene de resistência a ampicilina O plasmídeo obtido (pCYpp-slip1), contendo o gene sintético da lipase madura, fundiu a sequência líder prepo-fator-α, foi digerido (BamHI/bluntHindIII), e o fragmento resultante foi ligado dentro de pPICZαB linearizado com XbaI/blunt-HindIII, gerando pPIC-pp-slip1. pCY-pp-fator-α contém os fragmentos: SphI, BglI + gene sintétido da lipase contém os fragmentos: SphI, BglI + gene sintétido da lipase fragmento resultante ligado em pPICZαB pPIC-pp-slip1 Sequência líder prepo-fator--α linearizado com XbaI/blunt-HindIII
 Sequência que codifica a lipase. Sequência que codifica a lipase. Sequência que codifica a lipase. primer complementar. a sequência pPICZαB. O produto obtido da PCR foi clonado em SphI/blunt linearizado com pCYTEXPI, resultando em pCY-prepo-fator-α. O fragmento BglI de pCY-slip1, contendo toda a forma madura do gene sintético da lipase, foi inserido no plasmídeo digerido (pCyprepo-fator-α), restaurando assim o gene de resistência a ampicilina. O plasmídeo obtido (pCYpp-slip1), contendo o gene sintético da lipase madura, fundiu a sequência líder. prepo-fator-α, foi digerido (BamHI/bluntHindIII), e o fragmento resultante foi ligado dentro de. pPICZαB linearizado com XbaI/blunt-HindIII, gerando pPIC-pp-slip1. pCY-pp-fator-α. contém os fragmentos: SphI, BglI + gene sintétido da lipase. contém os fragmentos: SphI, BglI + gene sintétido da lipase. fragmento resultante ligado em pPICZαB. pPIC-pp-slip1. Sequência líder. prepo-fator--α. linearizado com XbaI/blunt-HindIII.")
46
Construção de vetores de expressão de Lip1
Como vetor de expressão em células de S. cerevisiae, foi usado um vetor contendo um promotor forte de indução da galactose (GALI-GALIO). pPIC-nl-slip1 foi digerido com BamHI e ligado a pYES2 com a mesma enzima, e desfosforilado, originando PY-nl-slip1. PY-nl-slip1 foi utilizado para clonagem no vetor pYES2 do gene sintético da lipase, procedido pelas sequências prepo-sinal fator alfa e pre-sinal. Os fragmentos HindIII-BsteII contendo os genes recombinantes foram isolados a partir de pPIC-pp-slip1 e pPIC-p-slip1, e inseridos em pYES2, digerido com a mesma enzima. Os plasmídeos obtidos foram chamados de pY-pp-slip1 e pY-p-slip1, respectivamente.
. pPIC-nl-slip1 foi digerido com BamHI e ligado a pYES2 com a mesma enzima, e desfosforilado, originando PY-nl-slip1. PY-nl-slip1 foi utilizado para clonagem no vetor pYES2 do gene sintético da lipase, procedido pelas sequências prepo-sinal fator alfa e pre-sinal. Os fragmentos HindIII-BsteII contendo os genes recombinantes foram isolados a partir de pPIC-pp-slip1 e pPIC-p-slip1, e inseridos em pYES2, digerido com a mesma enzima. Os plasmídeos obtidos foram chamados de pY-pp-slip1 e pY-p-slip1, respectivamente.")
47
Expressão de Lip1 em: P. pastoris S. cerevisiae

48
Expressão de Lip1 em P. pastoris
Cepa: GS115 Transformadas com o vetor pPIC-pp-slip1 por eletroporação Transformantes → plaqueamento em meio seletivo sólido (YEPD) contendo zeocina Clones produtores de lipase → halo claro Todos os transformantes obtidos com pPIC-pp-slip1 exibiram atividade lipásica após incubação 'overnight' detecção de microorg. lipolíticos verificação da atividade da lipase transferência das colônias para placas de tributirina-metanol incubadas a 30ºC 48h 0,1mL de metanol → adicionado a cada 24h em cada tampa células de Pichia transformadas com pPICαB nunca formaram halos
 contendo zeocina. Clones produtores de lipase → halo claro. Todos os transformantes obtidos com pPIC-pp-slip1 exibiram atividade lipásica após incubação overnight detecção de microorg. lipolíticos. verificação da atividade da lipase. transferência das colônias. para placas de tributirina-metanol. incubadas a 30ºC. 48h. 0,1mL de metanol → adicionado a cada 24h em cada tampa. células de Pichia transformadas com pPICαB nunca formaram halos.")
49
Expressão de Lip1 em S. cerevisiae
Células de S. cerevisiae Invsc2 Transformadas com pY-nl-slip1, pY-pp-slip1 e pYpreLIP por eletroporação Transformantes foram plaqueados em meio mínimo sólido contendo tributirina e galactose selecionados com base na capacidade de crescimento na ausência de leucina selecionados pela produção e secreção de lipase halos transparentes na placa contendo tributirina

50
adição de metanol após a cultura alcançar
Fermentações Realizada em um biorreator 1-L a 30ºC em meio padrão rico BMMY Fermentação com alta densidade de células Biorreatores: inoculados com 50mL da cultura de balão crescida 'overnight', com uma DO600=2-3 no meio BMGY mantida em pH constante Monitoramento: atividade lipolítica dos sobrenadantes, peso úmido das células pH 6,0 pH 7,0 adição de metanol após a cultura alcançar 75mg/mL de peso úmido adição de 5mL de metanol diariamente adição de HCl 2M, NaOH 2M

51
Caracterização físico-química e catalítica
Realizada diretamente com o sobrenadante da cultura, sem qualquer tipo de purificação medido rotineiramente com tributirina como substrato em um ensaio de pH-stat a 30ºC e pH 7,2 Atividade da lipase Atividade da lipase Atividade da lipase Atividade da lipase sobrenadante da cultura de pequena escala: ensaio com espectrotofômetro usando p-nitrofenil palmitato como substrato a 30ºC utilizou-se como substrato: 20mM de triglicérides 5% (w/v) de manteiga de cacau emulsificado em água destilada contendo goma arábica (20mg/mL) e usada como uma solução de substrato no ensaio de pH-stat a 30ºC e pH 7,2 Especificidade do substrato
 de manteiga de cacau emulsificado em água. destilada contendo goma arábica (20mg/mL) e usada como uma. solução de substrato no ensaio de pH-stat a 30ºC e pH 7,2. Especificidade. do substrato.")
52
Caracterização físico-química e catalítica
Emulsão triesterarina obtida pela adição de 5% de acetona (v/v) Retirou-se 2mL da mistura de reação, parando a reação lipolítica pela adição de 0,1mL de ácido ortofosfórico Foi extraído 3 vezes ácidos graxos livres com dietileter-n-hexano (1:1) e 50uL do extrato de ácidos graxos, que secou-se sob nitrogênio. nessas condições as lipases retêm 47% de sua plena atividade em tributirina 10mM de cada éster Especificidade do substrato Substrato: esteres metílicos de ácido graxo Preparou-se: 110mM da mistura dos esteres metílicos de ácido graxo e 5mM Tris-tampão com pH 7,5 Lipólise pH-stat a 30ºC e pH 7,2, até a titulação de 6,6mM de ácidos graxos livres
 Retirou-se 2mL da mistura de reação, parando a reação lipolítica pela adição de 0,1mL de ácido ortofosfórico. Foi extraído 3 vezes ácidos graxos livres com dietileter-n-hexano (1:1) e 50uL do extrato de ácidos graxos, que secou-se sob nitrogênio. nessas condições as lipases retêm. 47% de sua plena atividade em tributirina. 10mM de cada éster. Especificidade. do substrato. Substrato: esteres metílicos de ácido graxo. Preparou-se: 110mM da mistura dos esteres metílicos. de ácido graxo e 5mM Tris-tampão com pH 7,5. Lipólise. pH-stat a 30ºC e pH 7,2, até a titulação de 6,6mM de ácidos graxos livres.")
53
Características físico-química e catalítica
Os 3 ácidos graxos livres foram derivados para os silil éteres pela adição de 50uL de MSHFBA(N-metil-N-trimetil-hepta-fluorbutirâmido) e incubação a temperatura ambiente por 15 min Determinou-se pela cromatografia gasosa após uma nova adição de 50uL de etanol e 200uL de n-hexano (cromatografia gasosa: Fison 800), temperatura de 40ºC por 2 min, 4ºC por 1 min, 250ºC por 15 min, injetor a 350ºC, detector de ionização de chama a 360ºC a 75kPa
 e incubação a temperatura ambiente por 15 min. Determinou-se pela cromatografia gasosa após uma nova adição de 50uL de etanol e 200uL de n-hexano (cromatografia gasosa: Fison 800), temperatura de 40ºC por 2 min, 4ºC por 1 min, 250ºC por 15 min, injetor a 350ºC, detector de ionização de chama a 360ºC a 75kPa.")
54
Caracterização físico-química e catalítica
Efeito do pH Determinado a 30ºC e pH 7,2 Efeito da temperatura Determinado em diversas temperaturas Após a incubação das alíquotas da solução de lipase por : 20h a 30ºC em 0,1M de tampão de fosfato em diferentes pHs 30 min em 25 mM tampão Tris em pH 7,5 a diferentes temp Atividade da enzima

55
Métodos analíticos Sobrenadante que continha a lipase
Após 'blotting' em uma membrana PVDF, a banda da lipase foi cortada e utilizada para determinação da sequência N-terminal SDS-PAGE Análise da sequência N-terminal sequenciador de fase gasosa 470A (Applied Biosystems) Immunoblot Utilizando anticorpos policlonais específicos para a lipase homóloga de Geotrichum candidum
 Immunoblot. Utilizando anticorpos policlonais. específicos. para a lipase homóloga. de Geotrichum candidum.")
56
Métodos analíticos Deglicosilação das amostras
enzima relizada com endo-B-N-acetilglicosamindase H por 12h a 37ºC em um tampão de acetato de potássio de 50mM com pH 5,5 contendo 0,5mM de PMSF a proteína foi primeiramente incubada a 95ºC por 3 min com 0,1% (m/v) de SDS, e a reação foi realizada por 24h inibidor de proteases Deglicosilação das amostras desnaturadas
 de SDS, e a reação foi realizada por 24h. inibidor de proteases. Deglicosilação das amostras. desnaturadas.")
57
Obrigado!

Apresentações semelhantes
 dos genes e seu armazenamento>")


















