Carregar apresentação
A apresentação está carregando. Por favor, espere

1
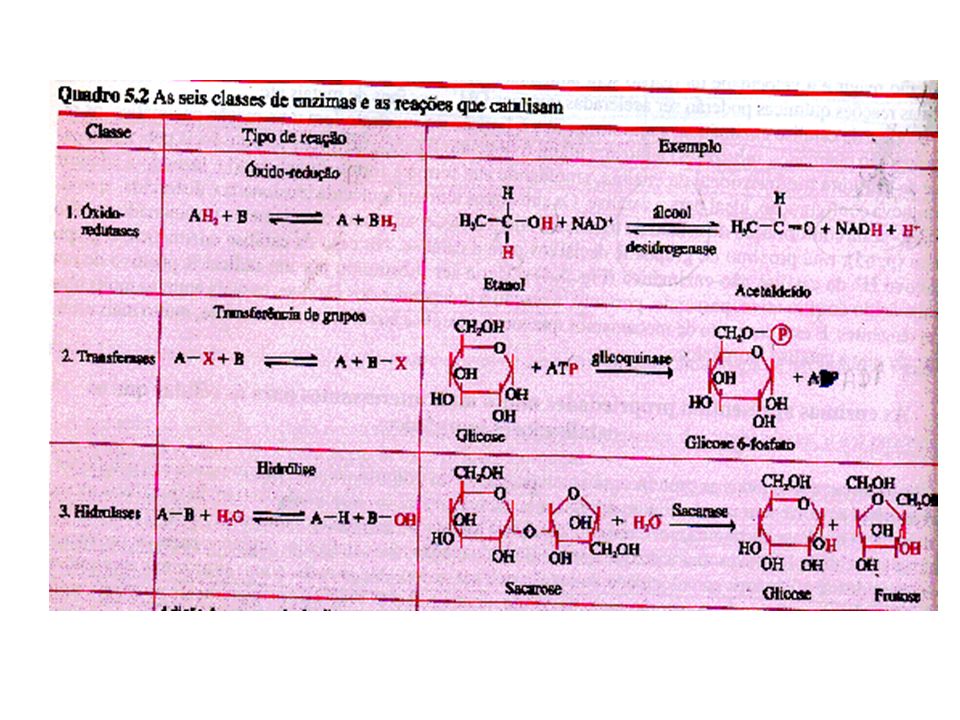
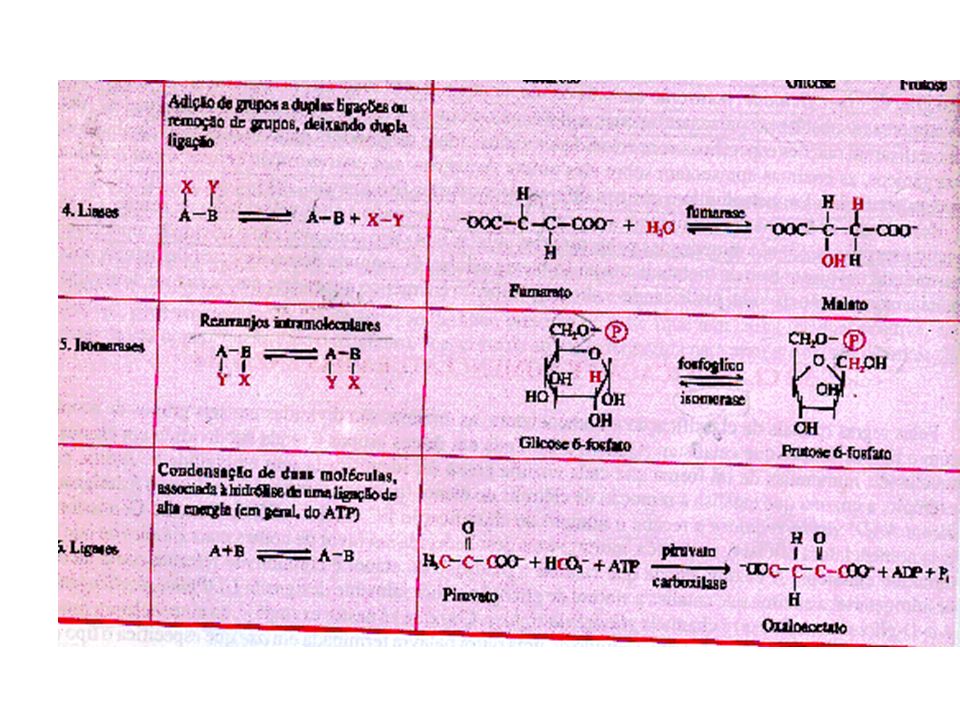
Introdução a Enzimas Enzimas -são catalisadores biológicos de alta especificidade. Catalisar uma reação química é alterar a sua velocidade. A presença de enzimas nas reações celulares aumenta a velocidade da reação por serem altamente específicas. Propriedades gerais das enzimas: Velocidade das reações mais rápida - as reações catalisadas por enzimas são de 106 a 1012 vezes mais rápidas que as correspondentes não catalisadas. Condições de reações mais brandas – as reações catalisadas por enzimas ocorrem em temperaturas inferiores a 100 C e pH quase neutro. Maior especificidade da reação Capacidade de regulação –

4
Especificidade dos substratos.
Complementaridade geométrica. Em geral o sítio de ligação do substrato à enzimas consiste em um sulco na superfície da enzima, complementar ao formato do substrato. Complementaridade eletrônica. Além disso, os resíduos de aminoácidos que formam o sítio de ligação estão organizados de modo a formar interações de atração específicas com o substrato. A ligação do substrato na enzima se dá em uma pequena e bem definida região da enzima chamada de centro ativo (ou sítio ativo da enzima). O substrato deve ter a forma espacial adequada para se alojar-se no centro ativo da enzima. Com isto esta ligação permite uma especificidade para a catálise.
. O substrato deve ter a forma espacial adequada para se alojar-se no centro ativo da enzima. Com isto esta ligação permite uma especificidade para a catálise.")
5
Os fatores que interferem na atividade enzimática:
1- pH A maioria das enzimas apresentam um valor de pH para qual a atividade da enzima é máxima. A velocidade diminui a medida que o pH se afasta do valor ótimo, que é característico para cada enzima. Geralmente o pH é neutro. 2- Temperatura. A temperatura também interfere na atividade da enzima. Se aumentarmos muito a temperatura a enzima pode perder sua forma nativa que permite desempenhar sua função levando a um processo de desarranjo estrutural (perda da estrutura terciária) chamado de Desnaturação. A desnaturação provoca drásticas alterações conformacionais na molécula, acarretando na perda da catálise. Ex: acima de 50 a 55 C a maioria das proteínas globulares são desnaturadas. 3- Concentração do Substrato.
 chamado de Desnaturação. A desnaturação provoca drásticas alterações conformacionais na molécula, acarretando na perda da catálise. Ex: acima de 50 a 55 C a maioria das proteínas globulares são desnaturadas. 3- Concentração do Substrato.")
6
Co-fatores: Muitas enzimas necessitam de associação com co-fatores não protéicos (moléculas ou íons) para exercer seu papel catalítico. Os cofatores comumente encontrados incluem íons metálicos (Cu2+, Fe3+ ou Zn2+ etc) ou moléculas orgânicas, não protéicas, de complexidade variada, que recebem o nome de coenzimas, tal com o NAD+. As coenzimas são co-fatores que se assoxiam temporariamente com uma dada molécula enzimática, de maneira que elas funcionam como co-substratos. Outros co-fatores são conhecidos como grupo prostéticos, que estão permanentemente ligados a sua proteínas geralmente através de ligações covalentes. Ex: heme o citrocromo c é fortemente ligado a proteína por uma extensa rede de interações hidrofóbicas e pontes de hidrogênio junto a ligações covalentes entre o heme e regiões específicas da proteínas.
 ou moléculas orgânicas, não protéicas, de complexidade variada, que recebem o nome de coenzimas, tal com o NAD+. As coenzimas são co-fatores que se assoxiam temporariamente com uma dada molécula enzimática, de maneira que elas funcionam como co-substratos. Outros co-fatores são conhecidos como grupo prostéticos, que estão permanentemente ligados a sua proteínas geralmente através de ligações covalentes. Ex: heme o citrocromo c é fortemente ligado a proteína por uma extensa rede de interações hidrofóbicas e pontes de hidrogênio junto a ligações covalentes entre o heme e regiões específicas da proteínas.")
7
Apoenzima (inativa) + co-fator holoenzima (ativa)
O conjunto da enzima com o seu co-fator apropriado e cataliticamente ativo é o chamado de holoenzima. A proteína enzimaticamente inativa resultante da remoção do co-fator da holoenzima é chamada de apoenzima; portanto a apoenzima é a porção protéica da holoenzima. Apoenzima (inativa) + co-fator holoenzima (ativa)
 + co-fator holoenzima (ativa)")
8
INTERAÇÃO ENZIMA-SUBSTRATO
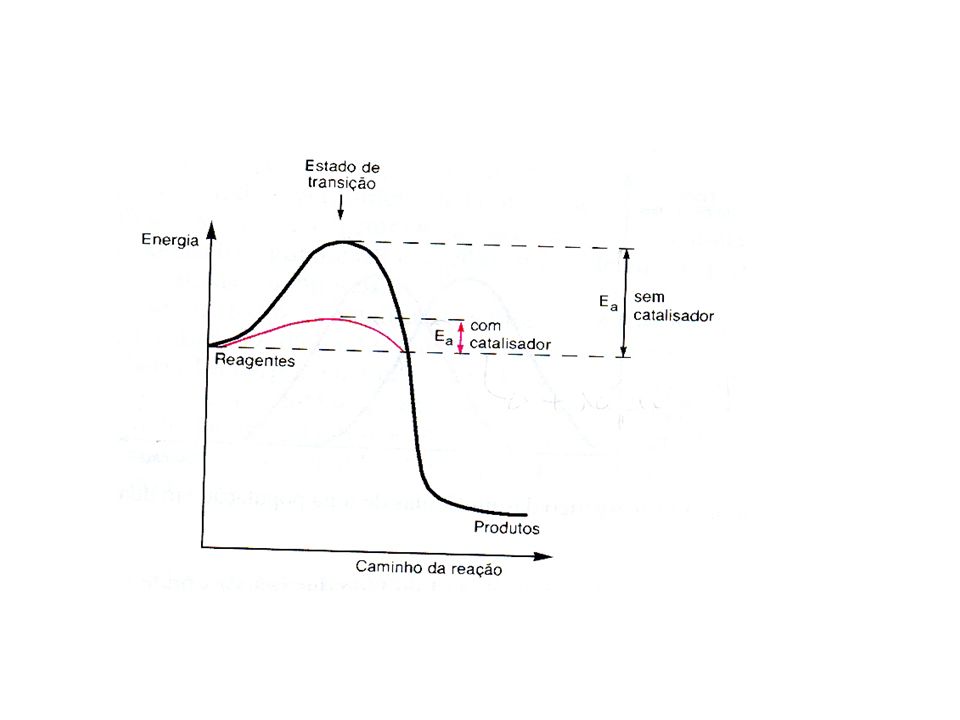
As enzimas aceleram a velocidade da reação por diminuir sua energia de ativação. A velocidade das reações é explicada pela teoria das colisões. Esta teria estabelece que, para a molécula reagir, as moléculas presentes em uma solução devem colidir com orientação apropriada e que a colisão deve levá-las a adquirir uma quantidade mínima de energia que lhes permita atingir o estado reativo, chamado de estado de transição. Para levar todas as moléculas de um mol até o estado de transição necessita uma quantidade de energia, chamada de energia de ativação. A energia de ativação é portanto a barreira que separa os reagentes do produtos. A velocidade de uma reação será diretamente proporcional ao número de moléculas com energia de ativação igual ou maior do que a energia do estado de transição.

10
Cinética da reação enzimática.
A + B C+ B temos que v = k [A] [B] O estudo das cinética das reações baseia-se na velocidade das reações, que é diretamente proporcional a concentração do reagente. Como a medida que se processa a reação a concentração do reagente diminui e portanto a velocidade também, passando a ser proporcional a nova concentração, estabeleceu-se a velocidade inicial (v0) proporcional a concentração inicial de A. A reação enzimática processa-se em duas etapas: A enzima (E) e substrato (S) formam um complexo transitório (ES) O produto é liberado (P) e a enzima volta na forma livre (E).
 proporcional a concentração inicial de A. A reação enzimática processa-se em duas etapas: A enzima (E) e substrato (S) formam um complexo transitório (ES) O produto é liberado (P) e a enzima volta na forma livre (E).")
11
E + S ES P + E k1 > k2 > k3 V1 = k1 [E] [s] V2 = k2 [ES]
Formação do complexo ES Formação da dissociação do complexo ES Formação do produto – etapa mais lenta e limitante k1 > k2 > k3
![E + S ES P + E k1 > k2 > k3 V1 = k1 [E] [s] V2 = k2 [ES]](http://slideplayer.com.br/slide/9787986/31/images/11/E+%2B+S+ES+%EF%82%AE+P+%2B+E+k1+%3E+k2+%3E+k3+V1+%3D+k1+%5BE%5D+%5Bs%5D+V2+%3D+k2+%5BES%5D.jpg "Formação do complexo ES. Formação da dissociação do complexo ES. Formação do produto – etapa mais lenta e limitante. k1 > k2 > k3.")
12
Os pressupostos acima foram estabelecidos por Michaellis e Menten, que com tratamento matemático desvendaram a cinética de um grupo de enzimas chamadas de enzimas michaelianas. As enzimas estão muito mais diluídas em solução do que o substrato e o números de moléculas de enzimas é muito inferior ao do substrato.

13
A análise das 3 situações de concentração diferentes de substrato na figura cima com o mesmo número de enzimas esclarece a influencia da concentração do substrato na velocidade da reação. Com maiores concentração de substrato, as velocidades de formação do Produto é cada vez maior, porque estará formando cada vez mais complexo ES. E + S ES 75% % 50% % 25% % 0% % Se a quantidade de substrato for muito grande, a quantidade, a concentração de E será praticamente nula, encontrando-se toda a enzima disponível sob a forma de ES.

14
Maior concentração possível do complexo ES é na situação onde há formação de 100% e a reação será processada na maior velocidade possível. Esta concentração é dita saturante e, a partir dela novas concentrações de substrato não terá efeito sobre a velocidade da reação, que atingiu seu valor máximo – Velocidade máxima (Vmax). Assim a velocidade da reação é sempre porpocional à concentração de ES. A velocidade inicial é obtida medindo-se a quantidade de produto formado em tempos suficientemente curtos para que no máximo 5% do substrato tenha sido transformado em produto.

15
Constante de Michaelis (Km)
Km é a concentração do substrato na qual a velocidade da reação corresponde a metade da velocidade máxima. Portanto uma enzima que tiver o menor valor de Km, ela atingirá a máxima eficiência catalítica em baixas concentrações do substrato. Ex: a hexoquinase pode aceitar a glicose ou a frutose como substrato. Mas o Km para a glicose é 0,15 mM e para a frutose é 1,5 mM. Isto significa que para a frutose ser substrato para a hexoquinase é necessário uma concentração de 10 vezes maior (1,5 mM) do que a glicose (0,15mM), portanto a hexoquinase tem uma maior afinidade pela glicose do que pela frutose Equação de Michaelis – Menten Vo = Vmáx [S] Km + [S]
 do que a glicose (0,15mM), portanto a hexoquinase tem uma maior afinidade pela glicose do que pela frutose. Equação de Michaelis – Menten. Vo = Vmáx [S] Km + [S]")
16
A Transformação da equação de Michaelis – Menten por Lineweaver-Burk facilitou a determinação do Km
1 = Km+ [S] v0 Vmax [S]

17
Inibidores da atividade enzimática.
Inibidores Irreversíveis – são aqueles que reagem geralmente através de ligações covalentes com as moléculas da enzimas, destruindo-as parcialmente. Um exemplo de inibidores irreversíveis é a penicilina seu inibidor liga-se especificamente a enzimas da via de síntese da parede bacteriana, inibindo-as, desprovidas de parede as células ficam sujeita a lise bacteriana.

18
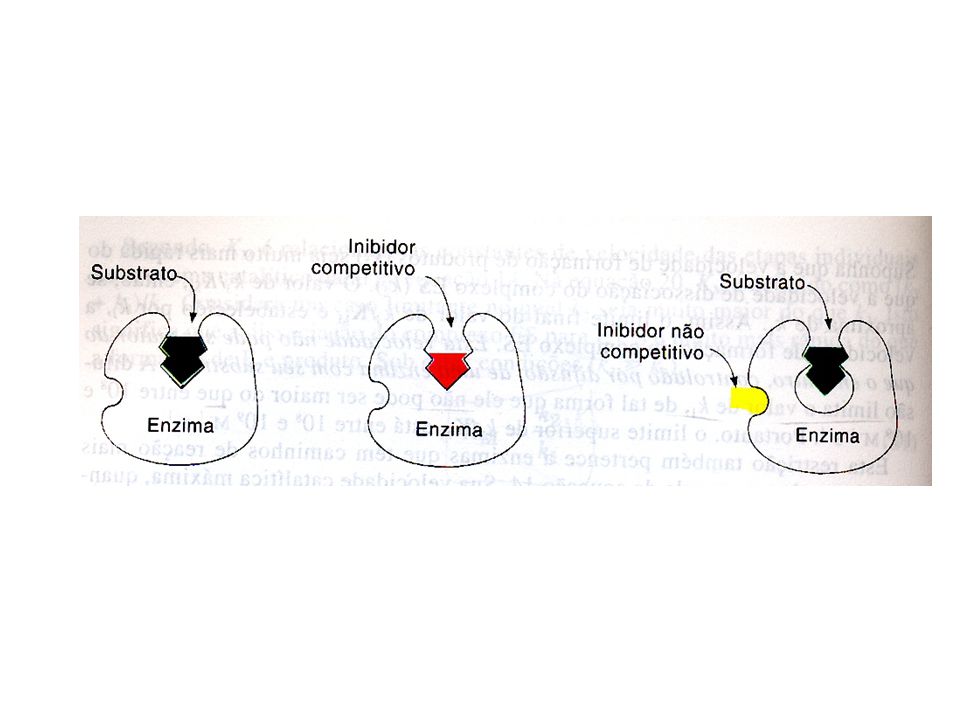
Inibidores reversíveis – são aqueles em que a inibição está relacionada com um equilíbrio entre enzima- inibidor. Podem ser competitivo e não competitivo. Inibição competitiva ( Ic )- ocorre quando o inibidor e o substrato tem estruturas semelhantes entre si e competem pelo centro ativo da enzima. Neste caso a velocidade máxima permanecerá inalterada e o valor de Km (afinidade da enzima pelo substrato) será aumentada. A velocidade máxima da reação será a mesma da reação efetuada sem a presença do inibidor, porem irá necessitar de uma maior concentração do substrato do que as reações sem a presença do inibidor, isto mostra que aparentemente há uma alteração no Km.
- ocorre quando o inibidor e o substrato tem estruturas semelhantes entre si e competem pelo centro ativo da enzima. Neste caso a velocidade máxima permanecerá inalterada e o valor de Km (afinidade da enzima pelo substrato) será aumentada. A velocidade máxima da reação será a mesma da reação efetuada sem a presença do inibidor, porem irá necessitar de uma maior concentração do substrato do que as reações sem a presença do inibidor, isto mostra que aparentemente há uma alteração no Km.")
19
Inibição não competitiva (INC) - o inibidor combina-se reversivelmente com a molécula da enzima em um outro ponto da estrutura, que não seja o seu centro ativo. Estes inibidores não são semelhantes estruturalmente à molécula do substrato. Neste caso a velocidade será alterada e o valor de Km (afinidade da enzima pelo substrato) permanecerá inalterado. E + INC E - INC . O valor de Km parece coincidir com o valor do novo km proporcionado pela diminuição da velocidade.

21
Inibição alostérica não obedecem à cinética de Mochaellis-Menten
As enzimas alostéricas apresentam normalmente 2 centros ativos: um para o substrato a ser transformado e outro para um inibidor que regula sua atividade, daí o seu nome. Geralmente as enzimas alostéricas possuem estrutura quaternária, e não seguem a cinética de Michaelis-Menten, mostrando-se uma curva sigmóide. Nas enzimas alostéricas, a ligação do substrato a um centro ativo pode afetar as propriedades dos outros centros ativos na mesma molécula de enzima. A atividade destas enzimas podem ser reguladas por inibidores, que se ligam a locais que não são os centros catalíticos. Ex: A ligação do oxigênio à hemoglobina é afetada por H+ e CO2.

Apresentações semelhantes