Carregar apresentação
A apresentação está carregando. Por favor, espere

1
Prof. Dr. Francisco Prosdocimi
Enzimologia Prof. Dr. Francisco Prosdocimi

2
A vida e a energia para a vida
Duas condições fundamentais: Autorreplicação Metabolismo Catálise enzimática A queima de açúcares é a principal forma segundo a qual retiramos energia do meio ambiente para vivermos Um saco de açúcar pode permane- cer anos na prateleira do supermercado A prateleira não tem enzimas! Nos animais, a glicose libera sua energia química em segundos Reações catalíticas promovem a oxidação da glicose ao quebrar ligações químicas que armazenam energia

3
Participam das vias bioquímicas
Enzimas realizam o controle preciso do metabolismo celular O metabolismo energético é um dos principais temas de estudo da bioquímica Permitem resposta e adaptação a um meio em mudança

4
O que são as enzimas? Proteínas notáveis, altamente especializadas
Alto grau de especificidade com substratos Poder catalítico extraordinário Muito maior do que catalisadores sintéticos Aceleram reações químicas Em condições suaves de temperatura e pH São o centro e o objeto de estudo principal da bioquímica Atuando de forma organizada catalisam centenas de reações que degradam as moléculas dos nutrientes e conservam suas energias Doenças ocorrem quando elas não funcionam bem Como os cientistas descobriram as enzimas?

5
LOUIS PASTEUR Um pouco de história...
1835: Berzelius, conceito de catálise 1885: fermentação do acúcar por lêvedos, gerando álcool Vitalismo: o mágico élan vital 1896: Edward Buchner consegue fermentar o açúcar num extrato de lêvedo sem vida! Fermentos, portanto, catalisavam reações químicas (açúcar a álcool) – biocatalisadores Enzima vem do grego εν ζυμη, cuja tradução é “no lêvedo” Louis Pasteur
 – biocatalisadores. Enzima vem do grego εν ζυμη, cuja tradução é no lêvedo Louis Pasteur")
6
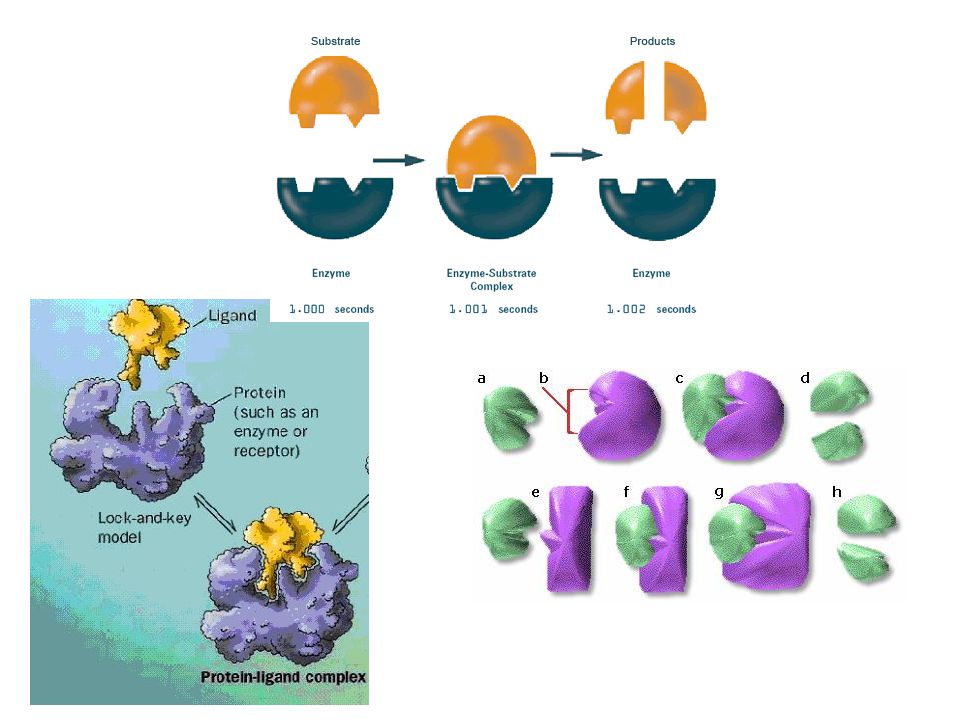
EMIL FISCHER Sacarase Quebra da sacarose em glicose e frutose Produziu diversos análogos de sacarose para testar se a enzima funcionava Determinadas mutações tornavam os análogos resistentes à sacarase Modelo de ação enzimática chave-e-fechadura Hermann Emil Fischer

8
James Batcheller Sumner
Enzimas são proteínas? Qual a natureza das enzimas? O químico orgânico alemão Richard Willstätter (1872–1942) – ganhador do Nobel pela estrutura da clorofila – conseguiu separar o componente enzimático de um preparado biológico e não encontrou nenhuma proteína! 1926 (EUA) – J Summer cristaliza a urease e conclui: enzimas são proteínas! Willstater criticou os resultados… James Batcheller Sumner
 – ganhador do Nobel pela estrutura da clorofila – conseguiu separar o componente enzimático de um preparado biológico e não encontrou nenhuma proteína! 1926 (EUA) – J Summer cristaliza a urease e conclui: enzimas são proteínas! Willstater criticou os resultados… James Batcheller Sumner")
9
SIM! Mas como funcionam? Kunitz e Northrop
Eletroforese e centrifugação: enzimas estão na fração protéica! Mesmo em quantidades proteícas indetectáveis pelos métodos, as enzimas continuavam tendo atividades Como as milhares de reações catalíticas eram possíveis a uma proteína? John Howard Northrop

10
? POLÍMEROS DE AMINOÁCIDOS?
1902: Emil Fischer (Estrasburgo) e Hofmeister (Berlim), proteínas eram formadas de aminoácidos que, ligados por ligações peptídicas, formavam cadeias polipeptídicas ?
 e Hofmeister (Berlim), proteínas eram formadas de aminoácidos que, ligados por ligações peptídicas, formavam cadeias polipeptídicas.")
11
Frederick Sanger 13 August 1918
Finalmente, Sanger 1952 Publica a primeira estrutura primária de uma proteína: a Insulina, com 51 aminoácidos O trabalho mostrava também que a estrutura das proteínas poderia ser descrita pela sua sequência de aminoácidos, do N ao C terminal A sequência, entretanto, não ajudava a prever a função da proteína (antes da bioinformática) Frederick Sanger 13 August 1918
 Frederick Sanger 13 August")
12
CONCLUSÃO: HISTÓRIA DA BIOQUÍMICA
As enzimas realizam reações catalíticas e transformam moléculas umas nas outras Os organismos biológicos são ricos em enzimas e as enzimas funcionam também fora dos organismos biológicos → biotecnologia! As enzimas são proteínas formadas por polímeros de aminoácidos A multiplicidade de função se dá pela interação tridimensional formada (modelo chave-fechadura) por interações não-covalentes a partir de uma série de aminoácidos ligados covalentemente (ligação peptídica) >gi|386828|gb|AAA | insulin [Homo sapiens] MALWMRLLPLLALLALWGPDPAAAFVNQHLCGSHLVEALYLVCGERGFFYTPKTR REAEDLQVGQVELGGGPGAGSLQPLALEGSLQKRGIVEQCCTSICSLYQLENYCN
 por interações não-covalentes a partir de uma série de aminoácidos ligados covalentemente (ligação peptídica) >gi|386828|gb|AAA | insulin [Homo sapiens] MALWMRLLPLLALLALWGPDPAAAFVNQHLCGSHLVEALYLVCGERGFFYTPKTR REAEDLQVGQVELGGGPGAGSLQPLALEGSLQKRGIVEQCCTSICSLYQLENYCN.")
13
Toda enzima é proteína?! Não!, há alguns RNAs que funcionam enquanto enzimas também Componente químico adicional necessário para a função Cofator: íons inorgânicos Coenzima: moléculas orgânicas complexas Se liga muito firmemente: grupo prostético Enzima completa: holoenzima Parte protéica: apoenzima ou apoproteína

14
Nomenclaturase das enzimases
Normalmente se adiciona o sufixo ase ao nome do substrato ou à atividade realizada Urease – hidrolisa a uréia DNA-polimerase – polimeriza DNA Pepsina – pepsis vem do grego (digestão) Sistema de classificação enzimático – EC number Quatro números: 2: transferase 7: fosfotransferase 1: transfere P para grupo OH- 1: tem D-glicose com aceptor
 Sistema de classificação enzimático – EC number. Quatro números: : transferase. 7: fosfotransferase. 1: transfere P para grupo OH- 1: tem D-glicose com aceptor.")
15
Reação enzimática Reação se dá em fases:
Enzima aumenta a velocidade das reações Os catalisadores aumentam a velocidade das reações por que diminuem a energia de ativação

16
Equilíbrio químico As enzimas realizam, muitas vezes, as reações nos dois sentidos A concentração de substratos e produtos é o que define a velocidade das reações Poder catalítico das enzimas vem da energia livre liberada na formação de ligações fracas quando da interação enzima-substrato Interações fracas entre ES são otimizadas no estado de transição

17
Bastonase Pauling (1946): Enzima deve ser complementar ao Estado de transição (ET), não ao substrato ET não é forma estável Interações fracas entre a enzima e o substrato propulsionam a catálise enzimática Necessidade de múltiplas interações fracas explica pq alguns prots são tão grandes Estabiliza o substrato

18
Especificidade enzimática
Deriva da formação de múltiplas interações fracas entre a enzima e a molécula do substrato específico Redução da entropia pela ligação Dessolvatação do substrato Ajuste induzido, proteína tbm muda de conformação

19
Grupos catalíticos Catálise geral ácido-base Catálise covalente
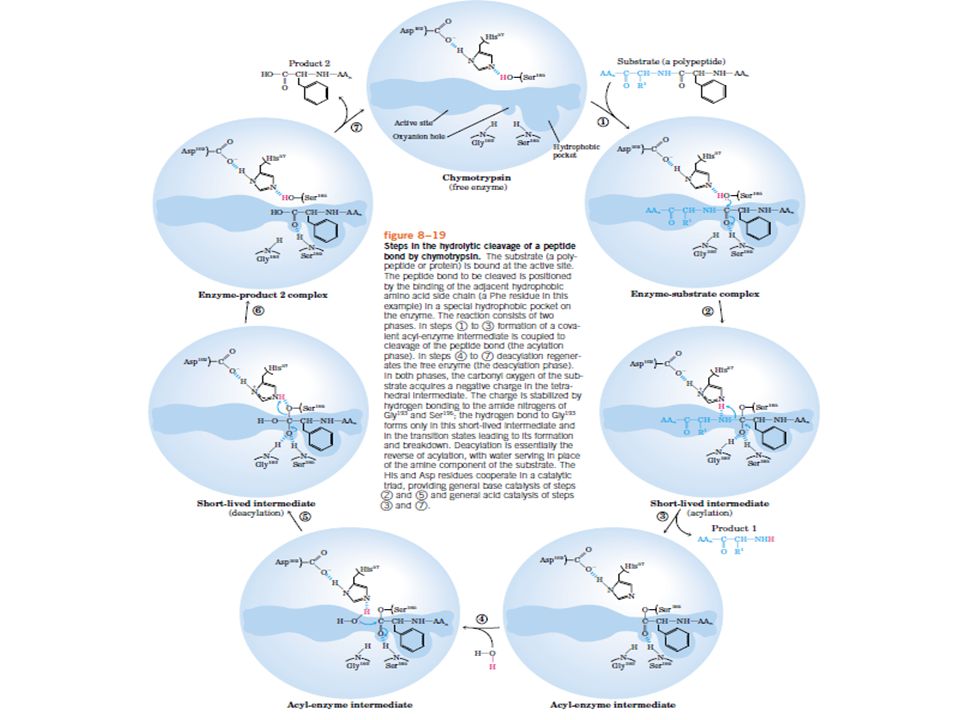
Transferência de prótons Catálise covalente Formação de lig. covalente transitória entre E e S Ativação do substrato Catálise por íons metálicos Estabilizam estados de transição 1/3 das enzimas conhecidas usam íons metálicos nas reações catalíticas Enzimas muitas vezes usam as três estratégias de catálise em conjunto -- quimiotripsina

20
Cinética enzimática Experimentos de mutagênese sítio-dirigida permite que os pesquisadores investiguem o papel de cada aminoácido na função protéica A concentração do substrato [S] influi na velocidade das reações catalisadas por enzimas Velocidade máxima é abstraída para concentrações excessivas de Substrato Constante de Michaelis kM = [S] correspondente a ½ Vmax;

21
Reação enzimática ocorre por etapas
Ligação reversível da enzima ao substrato Estado estacionário: concentrações constantes de enzima e enzima + substrato

22
Reações com 2 ou mais substratos
Enzimas podem formar os chamados complexos ternários ou realizar as reações uma-depois-da-outra Velocidade das reações químicas depende também da faixa de pH Maior velocidade está normalmente associada ao pH do ambiente onde a enzima atua

23
Inibição reversível Inibição competitiva
Inibidor compete pela ligação ao sítio ativo A ligação do inibidor altera os parâmetros cinéticos, tornando a reação mais lenta Os inibidores irreversíveis ligam-se covalentemente ou destroem grupos funcionais da enzima Podem ser usados como drogas

24
Exemplos de reações enzimáticas
Quimiotripsina Lisozima Hexocinase Enolase Beta-lactamases Proteases do HIV

25
A quimiotripsina Catalisa a hidrólise de ligações peptídicas adjacentes a aminoácidos aromáticos (Trp, Phe, Tyr) Forma intermediário acil-enzima covalente

27
A hexocinase Sofre um ajuste induzido quando ligada ao substrato
Fosforila um resíduo de glicose Primeiro passo da via glicolítica Adição de Xilose “engana” a enzima e faz com que ela fosforile a água

28
A enolase Realiza desidratação reversível de 2-fosfoglicertato a fosfoenolpiruvato Dímero com 436aa Catálise geral ácido base + estabilização do estado de transição Interações estabilizam intermediário (enolato) Ligações de H com outros aa’s do sítio ativo contribuem para o mecanismo geral
 Ligações de H com outros aa’s do sítio ativo contribuem para o mecanismo geral.")
29
A lisozima Agente antibacteriano natural encontrado nas lágrimas e clara de ovo Peptideoglicano da parede de bactérias é seu substrato Constituinte da parede microbiana que protege da lise osmótica Enzima rompe ligação glicosídica Monômero com 129 aa’s Mecanismo específico de ação enzimática ainda é controverso “mesmo uma infinidade de experimentos não pode provar que algo esteja certo, mas um único experimento pode provar que está errado” Albert Einstein

30
Medicamentos Muitos são inibidores de enzimas são usados para tratar doenças; desde as dores de cabeça até a AIDS Penicilina (Alexander Fleming, 1928) Peptideoglicano: polímero de açúcares e D-aa’s da parede bacteriana Penicilina interfere na síntese do peptideo glicano ao mimetizar um segmento D-Ala-D-Ala Inibem reação de transpeptidase
 Peptideoglicano: polímero de açúcares e D-aa’s da parede bacteriana. Penicilina interfere na síntese do peptideo glicano ao mimetizar um segmento D-Ala-D-Ala. Inibem reação de transpeptidase.")
31
Os Beta-lactâmicos Penicilina: degradada no ambiente do estômago
Penicilina V: estável em meio ácido pode ser administrada em via oral Amoxilina: ampla faixa de ação; antibiótico β-lactâmico + prescrito O ataque da Ser à porção amida do anel leva a um produto acil-enzima que é praticamente irreversível Transpeptidase inativa! Bloqueia síntese da parede bacteriana Rompe-se devido à pressão osmótica

32
As Beta-lactamases Enzimas que quebram o anel beta-lactâmico das penicilinas Disseminaram-se em bactérias submetidas à pressão seletiva O mau uso dos antibióticos

33
Guerra contra as bactérias
Acido clavulânico: inativa irreversivelmente as Beta-lactamases Clavulin: amoxilina + ácido clavulânico Foram encontradas bactérias resistentes ao ácido clavulânico... Desenvolvimento e descoberta de novos antibióticos é uma indústria em crescimento

34
Os antivirais: (1) o que é um vírus?
Anatomia molecular viral Ácido nucléico envolto por uma capa protetora feita de proteínas (capsídeo) Uma única molécula de ácido nucléico de fita simples ou dupla (fita positiva ou negativa) Genomas compactos Contêm apenas as proteínas necessárias à replicação viral Genomas da ordem de Kb
 Uma única molécula de ácido nucléico de fita simples ou dupla (fita positiva ou negativa) Genomas compactos. Contêm apenas as proteínas necessárias à replicação viral. Genomas da ordem de Kb.")
35
Parasitismo intra-celular
O vírus não tem metabolismo próprio Não é um organismo de vida-livre Parasita intra-celular obrigatório Forma de cristal Sequestra a maquinaria molecular da célula Mecanismo moleculares de produção preferencial dos genes virais (fator sigma próprio e específico)
")
36
Variedades virais Vírus de DNA Vírus de RNA
Precisam chegar ao núcleo para serem primeiramente transcritos em RNA Vírus de RNA Podem atuar no citoplasma Vírus de RNA com transcrição reversa Acontece a transcrição reversa do RNA em DNA e o DNA é normalmente integrado ao genoma do hospedeiro

37
HIV SIDA: Síndrome da imunodeficiência humana Retrovírus (lentivirus)
Pandemia Matou 25 milhões de pessoas nos últimos 25 anos Retrovírus (lentivirus) 2 cópias de um RNA fita simples (fita +) Nove genes Capsídeo é formado por cerca de 2000 proteínas p24 0,6% da população humana está infectada
 2 cópias de um RNA fita simples (fita +) Nove genes. Capsídeo é formado por cerca de 2000 proteínas p24. 0,6% da população humana está infectada.")
38
AZT (azidotimidina) Mimetiza um nucleotídeo de timina
Azida no lugar de hidroxila Prejudica a ação da transcriptase reversa Atrasa o desenvolvimento dos vírus, ainda que, aos poucos o vírus se torne resistente Transcriptase reversa mutante passa a não reconhecer mais o AZT

39
Outros tratamentos Inibidores de protease Inibidores de integrase
Impedem a protease do HIV de clivar as proteínas produzindo componentes maduros Inibidores de integrase Impedem o vírus de integrar seu DNA no genoma do hospedeiro Inibidores de fusão Interferem nas proteínas gp120 e gp41 ou bloqueiam receptores celulares de ligação ao HIV Coquetel anti-AIDS dificulta a adaptação do vírus a várias frentes moleculares de ataque, diminui comprovadamente o número de vírus no sangue

40
Proteases do HIV O RNA do vírus é traduzido em grandes proteínas
Essas proteínas precisam ser hidrolisadas em proteínas individuais do capsídeo A protease do HIV é uma aspartil-protease 2 resíduos de Asp facilitam ataque da água à ligação peptídica

41
Inibidores de proteases
Drogas que formam complexos não-covalentes com a protease do HIV Ligam-se fortemente: praticamente irreversíveis Cadeia principal com grupo hidroxil próximo a grupo benzil Grupo hidroxil mimetiza a água Resto da estrutura encaixa nas fendas da enzima Estrutura planejada para ser análoga ao estado de transição Sucesso terapêutico Aumentou longevidade e qualidade de vida dos doentes

42
Enzimas regulatórias Possui atividade catalítica aumentada ou diminuída em resposta a certos sinais Ajustes na velocidade da via metabólica permite que as células se adaptem a condições em variação Tipos mais comuns Enzimas alostéricas (ligações reversíveis a compostos) Modificações covalentes Ativadas por remoção de segmentos peptídicos (irreversível) Subunidade regulatória é normalmente diferente da subunidade catalítica
 Modificações covalentes. Ativadas por remoção de segmentos peptídicos (irreversível) Subunidade regulatória é normalmente diferente da subunidade catalítica.")
43
Enzimas alostéricas Homotrópicas: o ativador é o próprio substrato (hemoglobina) Heterotrópicas: outro ativador Enzimas alostéricas Mais de um sítio regulatório Cada um específico para um regulador Aspartato-transcarbamoilase 12 cadeias polipeptídicas Azul: catalíticas Vermelho/Amarelo: regulatórias

44
Enzimas alostéricas são reguladoras
... de vias bioquímicas Normalmente são o primeiro passo da via “economiza” a execução das outras reações Único passo de regulação da via Inibição por retroalimentação: São inibidas pelo produto do último passo Inibição alostérica heterotrópica

45
Regulação por modificações covalentes
500 tipos diferentes Modificações pós-traducionais covalentes já foram descritos Proteínas inteiras podem ser adicionados como a ubiquitina e a sumo Ubiquitinilação marca proteína para degradação Sumoilação é encontrada em proteínas nucleares Mudanças substanciais afetam de forma significativa a função da enzima Fosforilação é a mudança mais comum Podem haver vários sítios fosforiláveis

46
Proteínas cinases e fosfatases
Cinases: adicionam grupo fosforil a resíduos de Ser, Thr ou Tyr Básicas: fosforilam resíduos de vizinhança básica Preferências por resíduos próximos a Pro Atómos de O2 do grupo fosforil podem fazer pontes de H com outras regiões da proteína Reestabiliza a proteína estruturalmente Atuam em cascatas de sinalização celular

47
Cinases sítio-específicas
Fosforilações são sítio-específicas Cada enzima reconhece uma ordem de aminoácidos e fosforila resíduos de S, T ou Y Sítio de reconhecimento Sítio de fosforilação

48
Fosforilações múltiplas
São possíveis e permitem controle requintado da regulação Glicogênio sintase Catalisa a união de glicoses para formar glicogênio Fosforila em vários sítios (ver figura)
")
49
Proteólise de precursores
Proteases não podem ser produzidas em estado ativo Do contrário destruiriam as proteínas celulares... Zimogênios são precursores das proteínases Só funcionam depois da ativação por proteínas Ativação irreversível Mecanismo de produção de pró-proteínas ou pró-enzimas

50
Conclusões A atividade das vias metabólicas é regulada pelo controle da atividade de certas enzimas O conhecimento do mecanismo de ação das enzimas permite desenvolver medicamentos que inibam essa ação A inibição de uma enzima pode ser reversível, competitiva Os principais mecanismos de catálise são: ácido-básica, covalente e por íons metálicos As enzimas são catalisadores eficazes, aumentando a velocidade de ocorrência de uma reação

51
Aula baseada no livro do Lehninger (Nelson e Cox)
Capítulo 6 Enzimas

Apresentações semelhantes